Waggle dance
Waggle dance is a term used in beekeeping and ethology for a particular figure-eight dance of the honey bee. By performing this dance, successful foragers can share information about the direction and distance to patches of flowers yielding nectar and pollen, to water sources, or to new nest-site locations with other members of the colony.[1][2]
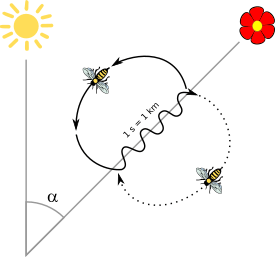
The waggle dance and the round dance are two forms of dance behaviour that are part of a continuous transition. As the distance between the resource and the hive increases, the round dance transforms into variations of a transitional dance, which, when communicating resources at even greater distances, becomes the waggle dance.[3] In the case of Apis mellifera ligustica, the round dance is performed until the resource is about 10 metres away from the hive, transitional dances are performed when the resource is at a distance of 20 to 30 metres away from the hive, and finally, when it is located at distances greater than 40 metres from the hive, the waggle dance is performed.[3] However, even close to the nest, the round dance can contain elements of the waggle dance, such as a waggle portion.[4] It has therefore been suggested that the term waggle dance is better for describing both the waggle dance and the round dance.[5]
Austrian ethologist and Nobel laureate Karl von Frisch was one of the first who translated the meaning of the waggle dance.[6]
Description

A waggle dance consists of one to 100 or more circuits, each of which consists of two phases: the waggle phase and the return phase. A worker bee's waggle dance involves running through a small figure-eight pattern: a waggle run (aka waggle phase) followed by a turn to the right to circle back to the starting point (aka return phase), another waggle run, followed by a turn and circle to the left, and so on in a regular alternation between right and left turns after waggle runs. Waggle-dancing bees produce and release two alkanes, tricosane and pentacosane, and two alkenes, (Z)-9-tricosene and (Z)-9-pentacosene, onto their abdomens and into the air.[7]
The direction and duration of waggle runs are closely correlated with the direction and distance of the resource being advertised by the dancing bee. In an experiment with capture and relocation of bees exposed to a waggle dance the bees followed the path that would have taken them to an experimental feeder had they not been displaced.[1] The resource can include the location of a food source or a potential nesting site.[8] For cavity-nesting honey bees, like the western honey bee (Apis mellifera) or Apis nigrocincta, flowers that are located directly in line with the sun are represented by waggle runs in an upward direction on the vertical combs, and any angle to the right or left of the sun is coded by a corresponding angle to the right or left of the upward direction. The distance between hive and recruitment target is encoded in the duration of the waggle runs.[1][9] The farther the target, the longer the waggle phase. The more excited the bee is about the location, the more rapidly it will waggle, so it will grab the attention of the observing bees, and try to convince them. If multiple bees are doing the waggle dance, it's a competition to convince the observing bees to follow their lead, and competing bees may even disrupt other bees' dances or fight each other off. In addition, some open-air nesting honeybees such as the black dwarf honeybee (Apis andreniformis), whose nests hang from twigs or branches, will perform a horizontal dance on a stage above their nest in order to signal to resources.[10]
Waggle dancing bees that have been in the nest for an extended time adjust the angles of their dances to accommodate the changing direction of the sun. Therefore, bees that follow the waggle run of the dance are still correctly led to the food source even though its angle relative to the sun has changed.
The consumption of ethanol by foraging bees has been shown to reduce waggle dance activity and increase occurrence of the tremble dance.[11]
Kevin Abbott and Reuven Dukas of McMaster University in Hamilton, Ontario, Canada discovered that if a dead western honeybee is placed on a flower, bees performed far fewer waggle dances upon returning to the hive. The scientists explain that the bees associate the dead bee with the presence of a predator at the food source. The reduction of the dance repetition frequency, therefore, indicates that the dancing bees perform and communicate a form of risk/benefit analysis.[12][13]
Though first decoded by Karl von Frisch, dancing behavior in bees had been observed and described multiple times prior. Around 100 years before Frisch's discovery, Nicholas Unhoch described dancing behavior of bees as being an indulgence "in certain pleasures and jollity".[6] He admitted ignorance as to the purpose of the dancing. 35 years prior to Unhoch's observations, Ernst Spitzner observed bees dancing and interpreted it as transmitting forage resource odors to other nestmates.[6] Aristotle, in addition to describing flower constancy behavior, suspected that some form of communication occurred between foragers within a nest:
"On each trip the bee does not fly from a flower of one kind to a flower of another, but flies from one violet, say, to another violet, and never meddles with another flower until it has got back to the hive; on reaching the hive they throw off their load, and each bee on her return is followed by three or four companions. What it is that they gather is hard to see, and how they do it has not been observed".[14]
Jürgen Tautz also writes about it in his book The Buzz about Bees (2008):
Many elements of the communication used to recruit miniswarms to feeding sites are also observed in "true" swarming behavior. Miniswarms of foragers are not placed under the same selection pressure as are true swarms, because the fate of the entire colony is not at stake. A truly swarming colony has to be quickly led to a new home, or it will perish. The behavior used to recruit to food sources possibly developed from the "true" swarming behavior.[15]
Mechanism
Honeybees accumulate an electric charge during flying and when their body parts are moved or rubbed together. Bees emit constant and modulated electric fields during the waggle dance. Both low- and high-frequency components emitted by dancing bees induce passive antennal movements in stationary bees according to Coulomb's Law. The electrically charged flagella of mechanoreceptor cells are moved by electric fields and more strongly so if sound and electric fields interact. Recordings from axons of the Johnston's organ indicate its sensitivity to electric fields. Therefore, it has been suggested that electric fields emanating from the surface charge of bees stimulate mechanoreceptors and may play a role in social communication during the waggle dance.[16]
Controversy

The dance language vs. the waggle dance
As defined by von Frisch, Tanzsprache (German for 'dance language') is the information about direction, distance, and quality of a resource (such as food or nesting sites) contained within the waggle dance.[17] There is supporting evidence of the waggle dance and "Tanzsprache" in Apis dorsata. Similar to other bees, they utilize the dance language to indicate the critical information regarding food resources. The dancer's body points in the direction of the food source and the sound produced during the dance indicates the profitability of the food.[18] Although there is some evidence for a direct connection between the Tanzsprache and the performance of the waggle dance, recent criticism holds that potential foragers need not correctly translate the dance language from the waggle dance to successfully forage.[17] In an experiment on the honeybee Apis mellifera, most individuals who thoroughly followed a waggle dance ignored the resource direction and location information. Instead, 93% of the foragers returned to foraging areas they had previous knowledge of.[17]
Bees that follow a waggle dance can successfully forage without decoding the dance language information in several ways:[19]
- Dance follower may use olfactory information from the dancer and find either the same resource or a different one with a similar scent.
- Following a dance may simply trigger foraging behavior. A forager may then search randomly for resources.
- Following a dance may reactivate private knowledge of a resource. After reactivation, the forager may return to the known resource.
- Using information communicated in the waggle dance is more useful to foragers when private information about resources is lacking.
Dance language as a language
The use of the word language may lead to misrepresentations of the waggle dance. The Swiss linguist Ferdinand de Saussure proposed a system of language where a sign is made up of two chief components. The signifier is the physical or phonetic representation of a sign. The signified is the conceptual component.[20] If the dance language followed the Saussurian dyadic model of semiotics, the signifier would be the waggle dance and the signified would be the location of the foraging resource. Though the dance language may or may not follow this sort of pattern, it is not considered to be a language with syntactical grammar or a set of symbols.[19]
Efficiency and adaptation
The waggle dance may be less efficient than once thought. Some bees observe over 50 waggle runs without successfully foraging, while others will forage successfully after observing 5 runs.[19] Likewise, studies have found that honeybees rarely make use of the information communicated in the waggle dance and seem to only do so about ten percent of the time.[21][22] Evidently there is a conflict between private information, or individual experience, and social information transmitted through dance communication. This sheds light on the fact that following social information is more energetically costly than foraging independently and is not always advantageous.[23][24] Using olfactory cues and memory of plentiful foraging sites, honeybees are able to successfully forage independently without expending the potentially extensive energy it takes to process and execute the directions communicated by their fellow foragers.
The waggle dance is beneficial in some environments and not in others, which provides a plausible explanation as to why the information provided by waggle dances are only used sparingly.[25] Depending on weather, other competitors, and food source characteristics, transmitted information may quickly degrade and become obsolete.[26][27] As a result, foragers have been reported to be attached to their food sites and continue to revisit a single patch many times after it has become unprofitable.[28] For example, the waggle dance plays a significantly larger role in foraging when food sources are not as abundant.[29][30] In temperate habitats, for instance, honey bee colonies routinely perform the waggle dance but were still able to successfully forage when the location information provided by the dance was experimentally obscured. In tropical habitats, however, honey bee foraging is severely impaired if waggle dancing is prevented. This difference is thought to be due to the patchiness of resources in tropical environment versus the homogeneity of resources in temperate environments. In the tropics, food resources can come in the form of flowering trees which are rich in nectar but are scattered sparsely and bloom only briefly. Thus, in tropical zones information about forage location might be more valuable than in temperate zones.[21]
Evolution
Ancestors to modern honeybees most likely performed excitatory movements to encourage other nest-mates to forage. These excitatory movements include shaking, zig-zagging, buzzing and crashing into nestmates. Similar behavior is observed in other Hymenoptera including stingless bees, wasps, bumblebees and ants.[19]
One promising theory for the evolution of the waggle dance, first proposed by Martin Lindauer, is that the waggle dance originally aided in the communication of information about a new nest site, rather than spatial information about foraging sites.[19][31]
Observations have suggested that different species of honeybees have different "dialects" of the waggle dance, each species or subspecies dance varying by curve or duration.[32][33] A study from 2008 demonstrated that a mixed colony of Asiatic honeybees (Apis cerana cerana) and European honeybees (Apis mellifera ligustica) were gradually able to understand one another's "dialects" of waggle dance.[34]
Applications to operations research
In line with recent work in swarm intelligence research involving optimization algorithms inspired by the behavior of social insects (including bees, ants and termites), and vertebrates such as fish and birds, there has recently been research on using bee waggle dance behavior for efficient fault-tolerant routing.[35] From the abstract of Wedde, Farooq, and Zhang (2004):[36]
In this paper we present a novel routing algorithm, BeeHive, which has been inspired by the communicative and evaluative methods and procedures of honey bees. In this algorithm, bee agents travel through network regions called foraging zones. On their way their information on the network state is delivered for updating the local routing tables. BeeHive is fault tolerant, scalable, and relies completely on local, or regional, information, respectively. We demonstrate through extensive simulations that BeeHive achieves a similar or better performance compared to state-of-the-art algorithms.
Another bee-inspired stigmergic computational technique called bee colony optimization is employed in Internet Server Optimization.[37][38]
The Zigbee RF protocol is named after the waggle dance.
References
- Riley JR, Greggers U, Smith AD, Reynolds DR, Menzel R (May 2005). "The flight paths of honeybees recruited by the waggle dance". Nature. 435 (7039): 205–7. Bibcode:2005Natur.435..205R. doi:10.1038/nature03526. PMID 15889092.
- Seeley TD, Visscher PK, Passino KM (2006). "Group decision making in honey bee swarms". American Scientist. 94 (3): 220–229. doi:10.1511/2006.3.220.
- Rinderer TE, Beaman LD (October 1995). "Genic control of honey bee dance language dialect". TAG. Theoretical and Applied Genetics. Theoretische und Angewandte Genetik. 91 (5): 727–32. doi:10.1007/bf00220950. PMID 24169907.
- Grüter C, Farina WM (May 2009). "The honeybee waggle dance: can we follow the steps?". Trends in Ecology & Evolution. 24 (5): 242–7. doi:10.1016/j.tree.2008.12.007. PMID 19307042.
- Gardner KE, Seeley TD, Calderone NW (2008-04-01). "Do honeybees have two discrete dances to advertise food sources?". Animal Behaviour. 75 (4): 1291–1300. doi:10.1016/j.anbehav.2007.09.032.
- von Frisch K (1967). The Dance Language and Orientation of Bees. Cambridge, Mass.: The Belknap Press of Harvard University Press.
- Thom C, Gilley DC, Hooper J, Esch HE (September 2007). "The scent of the waggle dance". PLoS Biology. 5 (9): e228. doi:10.1371/journal.pbio.0050228. PMC 1994260. PMID 17713987.
- "The Waggle Dance". www.pbs.org. Retrieved 2016-01-27.
- Radloff SE, Hepburn HR, Engel MS (2011). Honeybees of Asia. Berlin: Springer Science & Business Media. ISBN 978-3642164217.
- Raffiudin R, Crozier RH (May 2007). "Phylogenetic analysis of honey bee behavioral evolution". Molecular Phylogenetics and Evolution. 43 (2): 543–52. doi:10.1016/j.ympev.2006.10.013. PMID 17123837.
- Bozic J, Abramson CI, Bedencic M (April 2006). "Reduced ability of ethanol drinkers for social communication in honeybees (Apis mellifera carnica Poll.)". Alcohol. 38 (3): 179–83. doi:10.1016/j.alcohol.2006.01.005. PMID 16905444.
- Walker M (31 July 2009). "Honeybees warn of Risky Flowers". BBC Earth News. Retrieved 18 January 2010.
- Abbott KR, Dukas R (2009). "Honeybees consider flower danger in their waggle dance". Animal Behaviour. 78 (3): 633–635. doi:10.1016/j.anbehav.2009.05.029.
- Aristotle (1910). "Historia animalium, IX, 40, Becker 624b;". The Works of Aristotle. Translated by Thompson D. Clarendon, Oxford.
- Tautz J (2008). The Buzz about Bees - Biology of a Superorganism (photos by H. R. Heilmann). Heidelberg & Berlin: Springer. p. 112.
- Greggers U, Koch G, Schmidt V, Dürr A, Floriou-Servou A, Piepenbrock D, et al. (May 2013). "Reception and learning of electric fields in bees". Proceedings. Biological Sciences. 280 (1759): 20130528. doi:10.1098/rspb.2013.0528. PMC 3619523. PMID 23536603.
- Grüter C, Balbuena MS, Farina WM (June 2008). "Informational conflicts created by the waggle dance". Proceedings. Biological Sciences. 275 (1640): 1321–7. doi:10.1098/rspb.2008.0186. PMC 2602683. PMID 18331980.
- Kirchner W. "Acoustic signals in the dance language of the giant honeybee, Apis dorsata". Behavioral Ecology and Sociobiology. 33: 67–72. doi:10.1007/bf00171657.
- Grüter C, Farina WM (May 2009). "The honeybee waggle dance: can we follow the steps?". Trends in Ecology & Evolution. 24 (5): 242–7. doi:10.1016/j.tree.2008.12.007. PMID 19307042.
- de Saussure F (1916). "Nature of the Linguistics Sign". In Bally C, Sechehaye A (eds.). Cours de linguistique générale. McGraw Hill Education.
- Dornhaus A, Chittka L (2004). "Why do honey bees dance?". Behavioral Ecology and Sociobiology. 55 (4): 395–401. doi:10.1007/s00265-003-0726-9.
- Grüter C, Balbuena MS, Farina WM (June 2008). "Informational conflicts created by the waggle dance". Proceedings. Biological Sciences. 275 (1640): 1321–7. doi:10.1098/rspb.2008.0186. PMC 2602683. PMID 18331980.
- Al Toufailia H, Gruter C, Ratnieks FL (2013). "Persistence to unrewarding feeding locations by honeybee foragers (Apis mellifera): the effects of experience, resource profitability and season". Ethology. 119 (12): 1096–1106. doi:10.1111/eth.12170.
- Gruter C, Ratnieks FL (2011). "Honeybee foragers increase the use of waggle dance information when private information becomes unrewarding". Animal Behaviour. 81 (5): 949–954. doi:10.1016/j.anbehav.2011.01.014.
- Dornhaus A, Chittka L (2004-02-01). "Why do honey bees dance?". Behavioral Ecology and Sociobiology. 55 (4): 395–401. doi:10.1007/s00265-003-0726-9. ISSN 0340-5443.
- Grüter C, Farina WM (May 2009). "The honeybee waggle dance: can we follow the steps?". Trends in Ecology & Evolution. 24 (5): 242–7. doi:10.1016/j.tree.2008.12.007. PMID 19307042.
- Beekman M, Lew JB (March 2008). "Foraging in honeybees—when does it pay to dance?". Behavioral Ecology. 19 (2): 255–261. doi:10.1093/beheco/arm117. ISSN 1465-7279.
- Al Toufailia H, Grüter C, Ratnieks FL (December 2013). Herberstein M (ed.). "Persistence to Unrewarding Feeding Locations by Honeybee Foragers ( Apis mellifera ): the Effects of Experience, Resource Profitability and Season". Ethology. 119 (12): 1096–1106. doi:10.1111/eth.12170.
- Dyer FC (January 2002). "The biology of the dance language". Annual Review of Entomology. 47 (1): 917–49. doi:10.1146/annurev.ento.47.091201.145306. PMID 11729095.
- Grüter C, Balbuena MS, Farina WM (June 2008). "Informational conflicts created by the waggle dance". Proceedings. Biological Sciences. 275 (1640): 1321–7. doi:10.1098/rspb.2008.0186. PMC 2602683. PMID 18331980.
- Lindauer M (1961). Communication Among Social Bees. Cambridge: Harvard University Press. ISBN 978-0674147850.
- Gould JL, Towne WF (1989). "On the Evolution of the Dance Language: Response to Dyer and Seeley". American Naturalist. 134 (1): 156–159. doi:10.1086/284972. JSTOR 2462282.
- Dyer FC, Seeley TD (1991). "Dance Dialects and Foraging Range in Three Asian Honey Bee Species". Behavioral Ecology and Sociobiology. 28 (4): 227–233. doi:10.1007/BF00175094. JSTOR 4600541.
- Su S, Cai F, Si A, Zhang S, Tautz J, Chen S (June 2008). Giurfa M (ed.). "East learns from West: Asiatic honeybees can understand dance language of European honeybees". PLOS ONE. 3 (6): e2365. Bibcode:2008PLoSO...3.2365S. doi:10.1371/journal.pone.0002365. PMC 2391287. PMID 18523550.
- Crina G, Ajith A (2006). "Stigmergic Optimization: Inspiration, Technologies and Perspectives". Studies in Computational Intelligence. Heidelberg: Springer Berlin. 31: 1–24. ISBN 978-3-540-34689-0.
- Wedde HF, Farooq M, Pannenbaecker T, Vogel B, Mueller C, Meth J, Jeruschkat R (2005). "BeeAdHoc". Proceedings of the 2005 conference on Genetic and evolutionary computation - GECCO '05. p. 153. doi:10.1145/1068009.1068034. ISBN 978-1595930101.
- Nakrani S, Tovey C (2004). "On Honey Bees and Dynamic Server Allocation in Internet Hosting Centers". Adaptive Behavior. 12 (3–4): 223–240. CiteSeerX 10.1.1.115.3457. doi:10.1177/105971230401200308.
- Tovey C (Spring 2004). "The Honey Bee Algorithm: A Biological Inspired Approach to Internet Server Optimization". Engineering Enterprise: 13–15.
Further reading
- Gould JL (August 1975). "Honey bee recruitment: the dance-language controversy". Science. 189 (4204): 685–93. Bibcode:1975Sci...189..685G. doi:10.1126/science.1154023. PMID 1154023.
- Riley JR, Greggers U, Smith AD, Reynolds DR, Menzel R (May 2005). "The flight paths of honeybees recruited by the waggle dance". Nature. 435 (7039): 205–7. Bibcode:2005Natur.435..205R. doi:10.1038/nature03526. PMID 15889092.
- Seeley TD (1995). The Wisdom of the Hive. Cambridge, MA: Harvard University Press.
- von Frisch K (1967). The Dance Language and Orientation of Bees. Cambridge, MA: Harvard University Press.
- Rendell L, Boyd R, Cownden D, Enquist M, Eriksson K, Feldman MW, et al. (April 2010). "Why copy others? Insights from the social learning strategies tournament". Science. 328 (5975): 208–13. Bibcode:2010Sci...328..208R. doi:10.1126/science.1184719. PMC 2989663. PMID 20378813.
- Gruter C, Ratnieks FL (2011). "Honeybee foragers increase the use of waggle dance information when private information becomes unrewarding". Animal Behaviour. 81 (5): 949–954. doi:10.1016/j.anbehav.2011.01.014.
- Dyer FC (2002). "The biology of the dance language". Annual Review of Entomology. 47: 917–49. doi:10.1146/annurev.ento.47.091201.145306. PMID 11729095.
- Beekman M, Golag RS, Even N, Wattanchiyingchareon W, Olroyd BP (2008). "Dance precision of Apis florae--clues to the evolution of honeybee dance language". Behavioral Ecology and Sociobiology. 62 (8): 1259–1265. doi:10.1007/s00265-008-0554-z.
- Al Toufalia HM (2012). Honey bee foraging: persistence to non-rewarding feeding locations and waggle dance communication. Doctoral Thesis, University of Sussexx (doctoral). University of Sussex.
- Wenner AM, Wells PH (1990). Anatomy of a Controversy: The Question of a "Language" Among Bees. Columbia University Press, New York. pp. 299–311.
External links
| Wikimedia Commons has media related to Waggle dance. |
- The Waggle Dance of the Honeybee, Georgia Tech College of Computing on YouTube
- Waggle Dance Infographic - VetSci
- Communication and Recruitment to Food Sources by Apis mellifera — USDA-ARS
- Honeybee Communication — Kimball's Biology Pages