Mycangium
The term mycangium (pl., mycangia) is used in biology for special structures on the body of an animal that are adapted for the transport of symbiotic fungi (usually in spore form). This is seen in many xylophagous insects (e.g. horntails and bark beetles), which apparently derive much of their nutrition from the digestion of various fungi that are growing amidst the wood fibers. In some cases, as in ambrosia beetles (Coleoptera: Curculionidae: Scolytinae and Platypodinae), the fungi are the sole food, and the excavations in the wood are simply to make a suitable microenvironment for the fungus to grow. In other cases (e.g., the southern pine beetle, Dendroctonus frontalis), wood tissue is the main food, and fungi weaken the defense response from the host plant.[1]

crassiusculus
Some species of phoretic mites that ride on the beetles, have their own type of mycangium, but for historical reasons, mite taxonomists use the term acarinarium. Apart from riding on the beetles, the mites live together with them in their burrows in the wood.[2][3]
Origin
These structures were first systematically described by Helene Francke-Grosmann at 1956.[4] Then Lekh R. Batra[5] coined the word mycangia:[6] modern Latin, from Greek myco 'fungus' + angeion 'vessel'.
Function
The most common function of mycangia is preserving and releasing symbiotic inoculum. Usually, the symbiotic inoculum in mycangia will benefit their vectors (typically insect or mites), helping them to adapt to the new environment or provide nutrients of the vectors themselves and their descendants.[7]
For example, the ambrosia beetle (Euwallacea fornicatus) carries the symbiotic fungus Fusarium. When the beetle bores a host plant, it releases the symbiotic fungus from its mycangium. The symbiotic fungus becomes a plant pathogen, acting to weaken the resistance of host plant.[8] In the meantime, the fungus grows quickly in the galleries as the main food of beetle.[8] The offspring of beetle become mature, they will fill their mycangia with symbiont and hunt for the new host plant.[9]
Therefore, mycangia play an important role in protecting the inoculum from degradation and contamination. The structures of mycangia always resemble a pouch or a container, with caps or a small opening that reduce the possibility of contaminants from outside.[4] How mycangia release their inoculum is still unknown.
Mycangia and symbiotic inoculum
Most of the inoculum in mycangia are fungi. The symbiotic inoculum of most bark and ambrosia beetles are fungi belonging to Ophiostomatales (Ascomycota: Sordariomycetidae) and Microascales (Ascomycota: Hypocreomycetidae).[7] Symbiotic fungi in mycangia of woodwasps are Amylostereaceae (Basidiomycota: Russulales).[10] Symbiotic fungi in mycangia of lizard beetles are yeast (Ascomycota: Saccharomycetales).[11] Symbiotic fungi in mycangia of ship-timber beetles are Endomyces (Ascomycota: Dipodascaceae).[12] Symbiotic fungi in mycangia of leaf-rolling weevils are Penicillium fungi (Ascomycota: Trichocomaceae).[13] In addition to the above primary symbiotic fungi, secondary fungi and some bacteria have been isolated from mycangia.[14]
Mycangia in insects
Mycangia in bark and ambrosia beetles
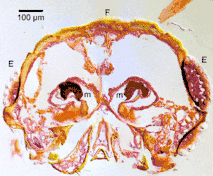
Mycangia of bark and ambrosia beetles (Curculionidae: Scolytinae and Platypodinae) are often complex cuticular invaginations for transport of symbiotic fungi.[2][7] Phloem-feeding bark beetles (Curculionidae: Scolytinae) have usually numerous small pits on the surface of their body, while ambrosia beetles (many Scolytinae and all Platypodinae), which are completely dependent on their fungal symbiont, have deep and complicated pouches.[7] These mycangia are often equipped with glands secreting substances to support fungal spores and perhaps to nourish mycelium during transport.[15] In many cases, the entrance to a mycangium is surrounded by tufts of setae, aiding in scraping mycelium and spores from walls of the tunnels and directing the spores into the mycangium. The mycangia of ambrosia beetle are highly diverse. Different genera or tribes with different kinds of mycangia. Some are oral mycangia in the head,[7] such as genus Ambrosiodmus and Euwallacea.[16] Some are pronotal mycangia, such genus Xylosandrus and Cnestus.[17]
Mycangia in woodwasps (horntails)
Mycangia of the woodwasps (Hymenoptera: Siricidae) were first described by Buchner.[18] Different from highly diverse types in bark and ambrosia beetles, woodwasps only have a pair of mycangia on the top of their ovipositor. Then when females deposit their eggs inside the host plant, they inject the symbiotic fungi from mycangia and phytotoxic mucus from another reservoir-like structure.[19]
Mycangia in lizard beetles
One species of lizard beetle Doubledaya bucculenta (Coleoptera: Erotylidae: Erotylidae) has mycangia on the tergum of the eighth abdominal segment. This ovipositor-associated mycangia is only present in adult females. Before Doubledaya bucculentnta deposit their eggs and inject the symbiotic microorganisms on a recently dead bamboo, they will excavate a small hole through the bamboo culm.[11]
Mycangia in ship-timber beetles
The ship-timber beetle (Coleoptera: Lymexylidae) is another family of wood-boring beetles that live with symbiotic fungi. Buchner first discovered their mycangia located on the ventral side of the long ovipositor.[20] These mycangia form a pair of integumental pouches at either side near the tip of oviduct. When the female lays the eggs, new eggs are coated with the fungal spores.
Mycangia in leaf-rolling weevils
Females of the leaf-rolling weevil in the genus Euops (Coleoptera: Attelabidae) store symbiotic fungi in the mycangia, which is between the first ventral segment of the abdomen and the thorax.[13] Different from ovipositor-associate mycangia in woodwasps, lizard beetles, and ship-timber beetles, mycangia of leaf-rolling weevils is a pair of spore incubators at the anterior end of the abdomen. This mycangium is formed by the coxa and the metendosternite at the posterior end of the thorax.[10]
Mycangia in stag beetles

Mycangia of the stag beetles (Coleoptera: Lucanidae) were discovered in Japan only this century [21]. This ovipositor-associated mycangium is located in a dorsal fold of the integument between the last two tergal plates of the adult females. It has been examined in many species [22][23][24]. A female everts the mycangium for the first time soon after eclosion; this is to retrieve the symbionts left by the larva on the pupal chamber when it emptied its gut before pupating. Later, when ovipositing, she everts it to pass on the inoculum to the next generation[25].
References
- Paine, T. D.; Stephen, F. M. (1987-01-01). "Fungi Associated with the Southern Pine Beetle: Avoidance of Induced Defense Response in Loblolly Pine". Oecologia. 74 (3): 377–379. Bibcode:1987Oecol..74..377P. doi:10.1007/bf00378933. JSTOR 4218483. PMID 28312476.
- Francke-Grossmann H. (1967). Ectosymbiosis in wood inhabiting insects. In: M. Henry (ed.) Symbiosis, Vol. 2. Academic Press, NewYork. pp.141-205.
- Mori, Boyd A.; Proctor, Heather C.; Walter, David E.; Evenden, Maya L. (2011-02-01). "Phoretic mite associates of mountain pine beetle at the leading edge of an infestation in northwestern Alberta, Canada". The Canadian Entomologist. 143 (1): 44–55. doi:10.4039/n10-043. ISSN 1918-3240.
- Francke-Grosmann, H. 1956. Hautdrüsen als träger der pilzsymbiose bei ambrosiakäfern. Zeitschrift für Morphologie und Ökologie der Tiere 45: 275–308.
- Batra, Lekh (1963). "Ecology of ambrosia fungi and their dissemination by beetles". Transactions of the Kansas Academy of Science. 66 (2): 213–236. doi:10.2307/3626562. JSTOR 3626562.
- Batra, L. R. (1963). "Ecology of ambrosia fungi and their dissemination by beetles". Trans. Kans. Acad. Sci. 66 (2): 213–236. doi:10.2307/3626562. JSTOR 3626562.
- Hulcr, Jiri; Stelinski, Lukasz L. (2017-01-31). "The Ambrosia Symbiosis: From Evolutionary Ecology to Practical Management". Annual Review of Entomology. 62 (1): 285–303. doi:10.1146/annurev-ento-031616-035105. PMID 27860522.
- Kasson, Matthew T.; O’Donnell, Kerry; Rooney, Alejandro P.; Sink, Stacy; Ploetz, Randy C.; Ploetz, Jill N.; Konkol, Joshua L.; Carrillo, Daniel; Freeman, Stanley (2013-07-01). "An inordinate fondness for Fusarium: Phylogenetic diversity of fusaria cultivated by ambrosia beetles in the genus Euwallacea on avocado and other plant hosts". Fungal Genetics and Biology. 56: 147–157. doi:10.1016/j.fgb.2013.04.004. PMID 23608321.
- "tea shot-hole borer, Euwallacea fornicatus". Featured Creatures.
- Sakurai, Kazuhiko (1985). "An attelabid weevil (Euops splendida) cultivates fungi". Journal of Ethology. 3 (2): 151–156. doi:10.1007/BF02350306. ISSN 0289-0771.
- Toki, Wataru; Tanahashi, Masahiko; Togashi, Katsumi; Fukatsu, Takema (2012-07-27). "Fungal Farming in a Non-Social Beetle". PLOS One. 7 (7): e41893. Bibcode:2012PLoSO...741893T. doi:10.1371/journal.pone.0041893. ISSN 1932-6203. PMC 3407107. PMID 22848648.
- Lyngnes, A. R. (1958). "Studier over Hylecoetus dermestoides L. under et angrep på bjorkestokker på Sunnmore 1954-1955". Norsk Entomologisk Tidsskrif. 10: 221–235.
- Kobayashi, Chisato; Fukasawa, Yu; Hirose, Dai; Kato, Makoto (2007-08-16). "Contribution of symbiotic mycangial fungi to larval nutrition of a leaf-rolling weevil". Evolutionary Ecology. 22 (6): 711–722. doi:10.1007/s10682-007-9196-2. ISSN 0269-7653.
- Hulcr, J.; Rountree, N. R.; Diamond, S. E.; Stelinski, L. L.; Fierer, N.; Dunn, R. R. (2012-05-01). "Mycangia of Ambrosia Beetles Host Communities of Bacteria". Microbial Ecology. 64 (3): 784–793. doi:10.1007/s00248-012-0055-5. ISSN 0095-3628. PMID 22546962.
- Six, Diana (2003). "Bark beetle-fungus symbioses". Insect Symbiosis. Contemporary Topics in Entomology. 20032558: 97–144. doi:10.1201/9780203009918.ch7. ISBN 978-0-8493-1286-1.
- Li, You; Simmons, David Rabern; Bateman, Craig C.; Short, Dylan P. G.; Kasson, Matthew T.; Rabaglia, Robert J.; Hulcr, Jiri (2015-09-14). "New Fungus-Insect Symbiosis: Culturing, Molecular, and Histological Methods Determine Saprophytic Polyporales Mutualists of Ambrosiodmus Ambrosia Beetles". PLOS One. 10 (9): e0137689. Bibcode:2015PLoSO..1037689Y. doi:10.1371/journal.pone.0137689. ISSN 1932-6203. PMC 4569427. PMID 26367271.
- Stone, W.D.; Nebeker, T.E.; Monroe, W.A.; MacGown, J.A. (2007-02-01). "Ultrastructure of the mesonotal mycangium of Xylosandrus mutilatus (Coleoptera: Curculionidae)". Canadian Journal of Zoology. 85 (2): 232–238. doi:10.1139/z06-205. ISSN 0008-4301.
- Buchner, P. 1928: Holznahrung und Symbiose. Vortrag gehalten auf dem X internationalen Zoologentag zu Budapest am 8. September 1927. Berlin: Springer, pp. 13–16.
- Coutts, M. P. (1969). "The mechanism of pathogenicity of Sirex noctilio in Pinus radiata. II. Effects of S. noctilio mucus". Aust. J. Biol. Sci. 22: 1153–1161. doi:10.1071/BI9691153.
- Casari, Sônia A.; Teixeira, Édson Possidônio (2011). "Larva of Atractocerus brasiliensis (Lepeletier & Audinet-Serville, 1825) (Lymexylidae, Atractocerinae)". Papéis Avulsos de Zoologia. 51 (12): 197–205. doi:10.1590/S0031-10492011001200001. ISSN 0031-1049.
- Tanahashi, M.; Kubota, K.; Matsushita, N.; Togashi, K. (2010). "Discovery of mycangia and the associated xylose-fermenting yeasts in stag beetles (Coleoptera: Lucanidae)". Naturwissenschaften. 97: 311–317.
- Tanahashi, M., Kubota, K., Matsushita, N.& Togashi, K. (2010). "Discovery of mycangia and the associated xylose-fermenting yeasts in stag beetles (Coleoptera: Lucanidae)". Naturwissenschaften. 97 (3): 311–317. Bibcode:2010NW.....97..311T. doi:10.1007/s00114-009-0643-5. PMID 20107974.CS1 maint: uses authors parameter (link)
- Tanahashi M., Fremlin M. (2013). "The mystery of the lesser stag beetle Dorcus parallelipipedus (L.) (Coleoptera: Lucanidae) mycangium yeasts". Bulletin of the Amateur Entomologists' Society. 72 (510): 146–152.
- Tanahashi, Masahiko; Hawes, Colin J. (2016). "The presence of a mycangium in European Sinodendron cylindricum(Coleoptera: Lucanidae) and the associated yeast symbionts". Journal of Insect Science. 16: 76. doi:10.1093/jisesa/iew054. PMC 4948600. PMID 27432353.
- Fremlin M., Tanahashi M. (2015). "Sexually-dimorphic post-eclosion behaviour in the European stag beetle Lucanus cervus (L.) (Coleoptera: Lucanidae)". Mitteilungen der Schweizerischen Entomologischen Gesellschaft. 88: 29–38.CS1 maint: uses authors parameter (link)