Strigolactone
Strigolactones are a group of chemical compounds produced by a plant's roots.[1] Due to their mechanism of action, these molecules have been classified as plant hormones or phytohormones.[1] So far, strigolactones have been identified to be responsible for three different physiological processes: First, they promote the germination of parasitic organisms that grow in the host plant's roots, such as Striga lutea and other plants of the genus Striga.[1] Second, strigolactones are fundamental for the recognition of the plant by symbiotic fungi, especially arbuscular mycorrhizal fungi, because they establish a mutualistic association with these plants, and provide phosphate and other soil nutrients.[1] Third, strigolactones have been identified as branching inhibition hormones in plants; when present, these compounds prevent excess bud growing in stem terminals, stopping the branching mechanism in plants.[1]
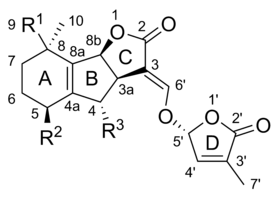
Strigolactones comprise a diverse group, but they all have core common chemical structure,[1] as shown in the image to the right. The structure is based on a tricyclic lactone linked to a hydroxymethyl butenolide; the former is represented in the figure as the A-B-C part, while the latter is the D part of the molecule.[1] It is important to note that most strigolactones present variations in the ABC part, but the D ring is quite constant across the different species, which led researchers to suspect that the biological activity relies on this part of the molecule.[1] Different studies have demonstrated that the activity of the molecules is lost when the C-D section of the molecules is modified.[1]
Since strigolactones are involved in the signaling pathway required for germination of parasitic species (such as Striga sp.), they have been a proposed target to control pests and overgrowth of these parasitic organism.[2] Using a molecule similar to strigolactones could be the key to designing a chemical and biological mechanism to stop the colonization of avplant's root by parasitic plants.[2]
Discovery
Germination of parasitic plant

Strigolactones were first isolated in 1966 from cotton plants, specifically from the roots. However its role in germination of other organisms was not determined until later.[3] Previous studies with Striga lutea had already shown that root extracts from the host plants were necessary for the parasitic seed to start germinating, which made obvious that a substance produced in the roots was stimulating this process.[3] The isolation of strigolactones lead to a series of tests that proved that this compound was the necessary molecule to induce germination of Striga species.[3] Later on, similar compounds were proven to produce the same effect: sorgolactone and alecrol, both of them presented the characteristic lactone group, so they were classified as strigolactones.[4] To induce germination of parasitic plants, strigolactones only needed to be present in trace amounts, in the order of 5 parts per million.[3]
Shoot branching hormone inhibition
The role of strigolactones as branching inhibitor hormone was discovered because of the use of a new set of mutant plants.[5] These mutants presented excessive growth in the axillary buds, which induced their terminal stem to start branching abnormally.[5] Previously, cytokinins were thought to be the only molecule involved in the regulation of stem branching, but these mutants presented normal production and signaling of cytokinins, leading to the conclusion that another substance was acting on the axillary buds.[5] Different tests that consisted in inserting part of the mutants plants into wild specimens (and vice versa), were able to demonstrated that the mutants were either not able to recognize a signal molecule coming from the roots and the lower part of the plant, or not able to produce the require molecules to inhibit branching.[5] This molecule, that was involved in branching regulation, was later identified to be a strigolactone.[5] The conclusion was that, in presence of strigolactones, the plant would be prevented to overgrowth and develop excessive branches, but when is not present, the axillary bud will start inducing abnormal branching.[5]
Chemistry
Properties
Although strigolactones vary in some of their functional groups, their melting point is usually found always between 200 and 202 Celsius degrees.[3] The decomposition of the molecule occurs after reaching 195oC.[3] They are highly soluble in polar solvents, such as acetone; soluble in benzene, and almost insoluble in hexane.[3]
Chemical structures
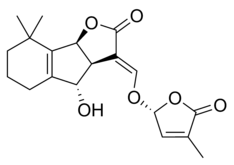
Some examples of strigolactones include:
-Strigol_chemical_structure.png) |
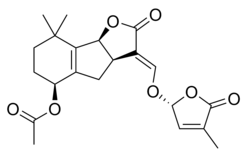 |
| (+)-Strigol | (+)-Strigyl acetate |
 |
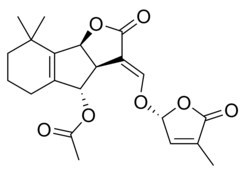 |
| (+)-Orobanchol | (+)-Orobanchyl acetate |
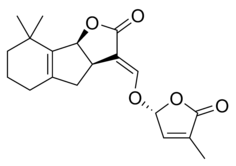 |
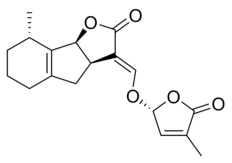 |
| (+)-5-Deoxystrigol | Sorgolactone |
Biosynthesis
Carotenoid pathway via carlactone
The biosynthetic pathway of the strigolactones has not been fully elucidated, but different steps have been identified, including the required enzymes to carry out the chemical transformation.[6] The first step is the isomerization of the 9th chemical bond of the -carotene, changing from trans configuration to cis.[6] This first step is carried out by the enzyme -carotene isomerase, also called DWARF27 or D27 for short, which required iron as a cofactor.[6] The second step is the chemical separation of 9-cis--carotene into two different compounds: the first one is 9-cis-aldehyde and the second is -ionone.[6] This second step is catalized by the carotenoid cleavage deoxygenase 7 (CCD7).[6] In the third step, another carotenoid cleavage oxygenase, called CCD8 (from the same family as CCD7), catalyze the conversion and rearrangement of the aldehyde created in the previous step into 9-cis--apo-10 and subsequently producing carlactone.[6]

It is still not clear how exactly carlactone is transformed into the different strigolactones identified so far, but several studies have proved that carlactone is definitely the precursor of strigolactones.[7] This last step of the biosynthesis should involve the addition of at least two oxygen molecules to convert the carlactone in 5-deoxystrigol, a simple strigolactone, and more oxidation should be required to produce other more complex strigolactone. The protein MAX1 has been proposed to catalyze the last step of the biosynthesis of strigolactones due its role in oxidative metabolism in plants.[7]
Role of ABA in biosynthesis
Both, abscisic acid (ABA) and strigolactones have a common group of enzymes that carried out the synthesis of the two compounds, previously it had been demonstrated the existence of a correlation of the two biosynthesis pathways, and it has been supported by different studies.[8][9] The ABA biosynthesis relies in a set of enzymes, called 9-cis-epoxycarotenoid dyoxygenase (NCED).[9] But, mutants plants that were defective in the production of the NCED enzymes, not just presented low levels of ABA, rather they also present low levels of strigolactones, specifically in the roots extracts where this hormone is mostly synthesized, this finding provided the basis for the existence of a common enzymatic machinery,[9] Other experiments that consist in blocking the NCED enzymes and using mutants unable to detect ABA changes, were used to support this theory.[8] So far there is a clear correlation of both synthesis that is related to the used of NCED enzymes in both biosynthesis, but the exact mechanism in which they are connected remains unclear.[8]
Molecular perception
In plants, strigolactones are perceived by the dual receptor/hydrolase protein DWARF14 (D14), a member of the α/β hydrolase superfamily. Despite being considered hydrolases with poor substrate turnover, an intact catalytic triad is required for the protein's biological function.[10] Several (in part competing) models have been proposed for the involvement of the catalytic triad in ligand perception:
- Hydrolysis of strigolactone, resulting in the D-ring being covalently attached to the active site serine.[11]
- Hydrolysis of strigolactone, resulting in a free D-ring that serves as a molecular glue at the entrance of the receptor, mediating interaction with another protein.[12]
- Binding of non-hydrolyzed, intact strigolactone that generates an altered DWARF14 protein surface, mediating interaction with another protein.[13]
- Hydrolysis of strigolactone, resulting in the D-ring being covalently attached to the active site histidine.[14][15][16][17]
- Hydrolysis of strigolactone, resulting in the D-ring being covalently attached to the active site serine and histidine at the same time, inducing a conformational change of DWARF14, leading to interaction with another protein.[18]
Kinetic results have suggested that the intact strigolactone triggers a signaling cascade after which hydrolysis is carried out as the final step to inactivate the strigolactone molecule.[19]
Mechanism of action
Germination of arbuscular mycorrhiza
Strigolactones are known to stimulate the germination of arbuscular mycorrhiza spores.[20] Since they produce this effect at extremely low concentrations, it has been proposed that the mechanism of activation must be a signaling pathway.[20] Different studies with diverse type of fungi, have found that after stimulation with strigolactones, the fungal cells present a higher amount of mitochondria and an increase in their oxidative activity.[20] Due the role of mitochondria in oxidative metabolism of macronutrients, it is thought that the spores remain inactive before finding the host plant, and once they are stimulated with strigolactones, the oxidative machinery in the mitochondrion gets activated to produce energy and nutrients necessaries for germination of the spore and fungal branching.[20] Studies with root extracts support this hypothesis, and so far strigolactones are the candidate molecules that better explain this increased in mitochondrial activity.[20]
Auxin-mediated secondary growth
It has been established that secondary growth in plant is mainly regulated by the phytohormone auxin.[21] However, the mechanism of auxin secretion is at the same time regulated by strigolactones, thus the latter can control secondary growth through auxin.[21] When applied in terminal buds of stem, strigolactone can block the expression of transport proteins required to move auxin across the buds, these proteins are denominated PIN1.[21] Thus, it was not surprising that when analyzing strigolactone deficient mutants, they were found to present an over-expression of PIN1 protein, which facilitate the transport of auxin in the terminal buds; auxin prevented the mitotic activity of these buds, stopping the plant to initiate secondary growth and branching.[21] In conclusion, plants depend in auxin transport for secondary growth initiation or inhibition, but these transport mechanism is dependent of the production of strigolactones, which can easily travel from the site of production (roots) to the terminal buds of the stem through the xylem.[21]
Ecology
Plant-fungi interaction

Strigolactones play a fundamental role in plant-fungi interaction.[22] One of the first studies made in Lotus japonicus had already demonstrated that compounds extracted from the root were necessary for the development of arbuscular mycorrhizal fungi that will establish a symbiotic relationship with the plant's root.[22] These same findings were true for different plants such as maize and sorghum.[22] Later on, the compounds responsible for the branching of the arbuscular fungi were isolated, and they include 5-deoxystrigol, strigol and sorgolactone, all of them belonging to the strigolactone family of compounds.[23][22] The process of branching is crucial to establish the symbiosis.[22] Since this branching only occurs after the germination of the spores and the initial growth of the hypha, the strigolactones required for germination have to be secreted by the plant and reached to fungi, meaning that strigolactones are also part of the recognition process by the fungi.[22]
Due the fact that arbuscula mychorriza can form symbiotic associations with the majority of the angiosperms, and many gymnosperms, it is expected to found different strigolactones compounds distributed in a whole variety of plants.[23] Unfortunately, while strigolactones are supposedly found in most plants, the studies done with strigolactones and AM fungi so far, have only studied a very limited range of plant species, mostly due the difficulty to extract these compounds and due its ease to disintegrate in solution.[23]
Strigolactones are not only necessary for the recognition of the plant by the fungi, they are also required by the recognition of the fungi by the plant.[24] The mechanism of fungal recognition occurs in a similar fashion that the recognition of bacterias such as Rhizobia sp.[24] In fact, it has been proposed that the recognition mechanism for bacterias evolved from the mechanism to recognize fungi, because the latter is known to be more primitive and ancient.[24] Just like bacteria use Nod factors, the fungi use a set of molecules denominated Myc factor.[24] These fungal products can be recognized by different plants and are not designed to be plant-specific.[24] When these Myc factors are recognized by the plant's root, they stimulate the expression of different genes involved in the initiation of the symbiotic association.[24] However, the secretion of the Myc factor by the fungi occurs only after being previously stimulated by strigolactones from the plant, demonstrating the necessary role of these compound for both recognition (from fungi and from plant).[24] Strigolactones also have been reported to produces other changes in fungal cells, such as an increase in the concentration of intracellular calcium and an increase in lipochitoolisaccharides (LCOs), the latter has been proved to be one of the Myc factors produced by the fungi for its recognition by the plant.[24]
One of the main roles of arbuscular fungi contained in symbiotic association with plants, is to provide soil nutrients to the plants, especially phosphate.[25] Thus when the phosphate in the depletion zone gets really low, the plant depend mainly in the AM fungi to fulfill its phosphate demands.[25] Studies with tomato plants have shown that, when plants undergo a deficit in phosphate, they produce higher amount of strigolactones, which in turn will increase the branching of AM fungi.[25] This excess development of the fungi is expected to provide the additional phosphate required for the plant, since the fungi can now spread to more soil areas.[25] However, since strigolactone also stimulate the germination of parasitic plants, these phosphate-deficient plants, also present higher invasion of parasitic species such as Striga sp.[25] Providing adequate phosphate through soil fertilization have been proved to reduce the proliferation of these parasites, because they require strigolactone for its germination.[25]
References
- Umehara M, Cao M, Akiyama K, Akatsu T, Seto Y, Hanada A, et al. (June 2015). "Structural Requirements of Strigolactones for Shoot Branching Inhibition in Rice and Arabidopsis". Plant & Cell Physiology. 56 (6): 1059–72. doi:10.1093/pcp/pcv028. PMID 25713176.
- Waters MT, Gutjahr C, Bennett T, Nelson DC (April 2017). "Strigolactone Signaling and Evolution". Annual Review of Plant Biology. 68 (1): 291–322. doi:10.1146/annurev-arplant-042916-040925. PMID 28125281.
- Cook CE, Whichard LP, Turner B, Wall ME, Egley GH (December 1966). "Germination of Witchweed (Striga lutea Lour.): Isolation and Properties of a Potent Stimulant". Science. 154 (3753): 1189–90. Bibcode:1966Sci...154.1189C. doi:10.1126/science.154.3753.1189. PMID 17780042.
- Xie X, Yoneyama K, Yoneyama K (2010-07-01). "The strigolactone story". Annual Review of Phytopathology. 48 (1): 93–117. doi:10.1146/annurev-phyto-073009-114453. PMID 20687831.
- Dun EA, Brewer PB, Beveridge CA (July 2009). "Strigolactones: discovery of the elusive shoot branching hormone". Trends in Plant Science. 14 (7): 364–72. doi:10.1016/j.tplants.2009.04.003. PMID 19540149.
- Alder A, Jamil M, Marzorati M, Bruno M, Vermathen M, Bigler P, et al. (March 2012). "The path from β-carotene to carlactone, a strigolactone-like plant hormone". Science. 335 (6074): 1348–51. Bibcode:2012Sci...335.1348A. doi:10.1126/science.1218094. PMID 22422982.
- Seto Y, Yamaguchi S (October 2014). "Strigolactone biosynthesis and perception". Current Opinion in Plant Biology. 21: 1–6. doi:10.1016/j.pbi.2014.06.001. PMID 24981923.
- Liu J, He H, Vitali M, Visentin I, Charnikhova T, Haider I, et al. (June 2015). "Osmotic stress represses strigolactone biosynthesis in Lotus japonicus roots: exploring the interaction between strigolactones and ABA under abiotic stress" (PDF). Planta. 241 (6): 1435–51. doi:10.1007/s00425-015-2266-8. hdl:2318/1508108. PMID 25716094.
- López-Ráez JA, Kohlen W, Charnikhova T, Mulder P, Undas AK, Sergeant MJ, et al. (July 2010). "Does abscisic acid affect strigolactone biosynthesis?" (PDF). The New Phytologist. 187 (2): 343–54. doi:10.1111/j.1469-8137.2010.03291.x. PMID 20487312.
- Hamiaux C, Drummond RS, Janssen BJ, Ledger SE, Cooney JM, Newcomb RD, Snowden KC (November 2012). "DAD2 is an α/β hydrolase likely to be involved in the perception of the plant branching hormone, strigolactone". Current Biology. 22 (21): 2032–6. doi:10.1016/j.cub.2012.08.007. PMID 22959345.
- Zhao LH, Zhou XE, Wu ZS, Yi W, Xu Y, Li S, et al. (March 2013). "Crystal structures of two phytohormone signal-transducing α/β hydrolases: karrikin-signaling KAI2 and strigolactone-signaling DWARF14". Cell Research. 23 (3): 436–9. doi:10.1038/cr.2013.19. PMC 3587710. PMID 23381136.
- Kagiyama M, Hirano Y, Mori T, Kim SY, Kyozuka J, Seto Y, et al. (February 2013). "Structures of D14 and D14L in the strigolactone and karrikin signaling pathways". Genes to Cells. 18 (2): 147–60. doi:10.1111/gtc.12025. PMID 23301669.
- Zhao LH, Zhou XE, Yi W, Wu Z, Liu Y, Kang Y, et al. (November 2015). "Destabilization of strigolactone receptor DWARF14 by binding of ligand and E3-ligase signaling effector DWARF3". Cell Research. 25 (11): 1219–36. doi:10.1038/cr.2015.122. PMC 4650425. PMID 26470846.
- Yao, Ruifeng; Ming, Zhenhua; Yan, Liming; Li, Suhua; Wang, Fei; Ma, Sui; Yu, Caiting; Yang, Mai; Chen, Li; Chen, Linhai; Li, Yuwen (25 August 2016). "DWARF14 is a non-canonical hormone receptor for strigolactone". Nature. 536 (7617): 469–473. Bibcode:2016Natur.536..469Y. doi:10.1038/nature19073. ISSN 1476-4687. PMID 27479325.
- de Saint Germain A, Clavé G, Badet-Denisot MA, Pillot JP, Cornu D, Le Caer JP, et al. (October 2016). "An histidine covalent receptor and butenolide complex mediates strigolactone perception". Nature Chemical Biology. 12 (10): 787–794. doi:10.1038/nchembio.2147. PMC 5030144. PMID 27479744.
- Bürger M, Mashiguchi K, Lee HJ, Nakano M, Takemoto K, Seto Y, et al. (January 2019). "Structural Basis of Karrikin and Non-natural Strigolactone Perception in Physcomitrella patens". Cell Reports. 26 (4): 855–865.e5. doi:10.1016/j.celrep.2019.01.003. PMID 30673608.
- Bürger, Marco; Chory, Joanne (April 2020). "The Many Models of Strigolactone Signaling". Trends in Plant Science. 25 (4): 395–405. doi:10.1016/j.tplants.2019.12.009. ISSN 1878-4372. PMID 31948791.
- Yao R, Ming Z, Yan L, Li S, Wang F, Ma S, et al. (August 2016). "DWARF14 is a non-canonical hormone receptor for strigolactone". Nature. 536 (7617): 469–73. Bibcode:2016Natur.536..469Y. doi:10.1038/nature19073. PMID 27479325.
- Seto Y, Yasui R, Kameoka H, Tamiru M, Cao M, Terauchi R, et al. (January 2019). "Strigolactone perception and deactivation by a hydrolase receptor DWARF14". Nature Communications. 10 (1): 191. Bibcode:2019NatCo..10..191S. doi:10.1038/s41467-018-08124-7. PMC 6331613. PMID 30643123.
- Besserer A, Puech-Pagès V, Kiefer P, Gomez-Roldan V, Jauneau A, Roy S, et al. (July 2006). "Strigolactones stimulate arbuscular mycorrhizal fungi by activating mitochondria". PLOS Biology. 4 (7): e226. doi:10.1371/journal.pbio.0040226. PMC 1481526. PMID 16787107.
- Shinohara N, Taylor C, Leyser O (2013-01-29). "Strigolactone can promote or inhibit shoot branching by triggering rapid depletion of the auxin efflux protein PIN1 from the plasma membrane". PLOS Biology. 11 (1): e1001474. doi:10.1371/journal.pbio.1001474. PMC 3558495. PMID 23382651.
- López-Ráez JA, Charnikhova T, Gómez-Roldán V, Matusova R, Kohlen W, De Vos R, et al. (2008-06-01). "Tomato strigolactones are derived from carotenoids and their biosynthesis is promoted by phosphate starvation". The New Phytologist. 178 (4): 863–74. doi:10.1111/j.1469-8137.2008.02406.x. hdl:10261/159227. PMID 18346111.
- Yoneyama K, Xie X, Sekimoto H, Takeuchi Y, Ogasawara S, Akiyama K, Hayashi H, Yoneyama K (July 2008). "Strigolactones, host recognition signals for root parasitic plants and arbuscular mycorrhizal fungi, from Fabaceae plants". The New Phytologist. 179 (2): 484–94. doi:10.1111/j.1469-8137.2008.02462.x. PMID 19086293.
- Genre A, Chabaud M, Balzergue C, Puech-Pagès V, Novero M, Rey T, et al. (April 2013). "Short-chain chitin oligomers from arbuscular mycorrhizal fungi trigger nuclear Ca2+ spiking in Medicago truncatula roots and their production is enhanced by strigolactone". The New Phytologist. 198 (1): 190–202. doi:10.1111/nph.12146. hdl:2318/134858. PMID 23384011.
- Bouwmeester HJ, Roux C, Lopez-Raez JA, Bécard G (May 2007). "Rhizosphere communication of plants, parasitic plants and AM fungi". Trends in Plant Science. 12 (5): 224–30. doi:10.1016/j.tplants.2007.03.009. PMID 17416544.