Mitochondrial ROS
Mitochondrial ROS (mtROS or mROS) are reactive oxygen species (ROS) that are produced by mitochondria.[1][2][3] Generation of mitochondrial ROS mainly takes place at the electron transport chain located on the inner mitochondrial membrane during the process of oxidative phosphorylation (OXPHOS). Leakage of electrons at complex I and complex III from electron transport chains leads to partial reduction of oxygen to form superoxide. Subsequently, superoxide is quickly dismutated to hydrogen peroxide by two dismutases including superoxide dismutase 2 (SOD2) in mitochondrial matrix and superoxide dismutase 1 (SOD1) in mitochondrial intermembrane space. Collectively, both superoxide and hydrogen peroxide generated in this process are considered as mitochondrial ROS.[1]
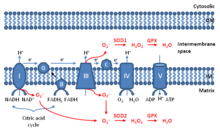
Once thought as merely the by-products of cellular metabolism, mitochondrial ROS are increasingly viewed as important signaling molecules,[4] whose levels of generation at 11 currently-identified sites vary depending on cellular energy supply and demand.[5][6] At low levels, mitochondrial ROS are considered to be important for metabolic adaptation as seen in hypoxia[1]. Mitochondrial ROS, stimulated by danger signals such as lysophosphatidylcholine and Toll-like receptor 4 and Toll-like receptor 2 bacterial ligands lipopolysaccharide (LPS) and lipopeptides, are involved in regulating inflammatory response[7][8]. Finally, high levels of mitochondrial ROS activate apoptosis/autophagy pathways capable of inducing cell death.[9]
COVID-19
Monocytes/macrophages are the most enriched immune cell types in the lungs of COVID-19 patients and appear to have a central role in the pathogenicity of the disease. These cells adapt their metabolism upon infection and become highly glycolytic, which facilitates SARS-CoV-2 replication. The infection triggers mitochondrial ROS production, which induces stabilization of hypoxia-inducible factor-1α (HIF1A) and consequently promotes glycolysis. HIF1A-induced changes in monocyte metabolism by SARS-CoV-2 infection directly inhibit T cell response and reduce epithelial cell survival. Targeting mitochondrial ROS may have great therapeutic potential for the development of novel drugs to treat patients with coronavirus.[10]
Aging
Mitochondrial ROS can promote cellular senescence and aging phenotypes in the skin of mice.[11] Ordinarily mitochondrial SOD2 protects against mitochondrial ROS. Epidermal cells in mutant mice with a genetic SOD2 deficiency undergo cellular senescence, nuclear DNA damage, and irreversible arrest of proliferation in a portion of their keratinocytes.[11][12]
Mutant mice with a conditional deficiency for mitochondrial SOD2 in connective tissue have an accelerated aging phenotype.[13] This aging phenotype includes weight loss, skin atrophy, kyphosis (curvature of the spine), osteoporosis, muscle degeneration and reduced life span.
DNA damage
Mitochondrial ROS attack DNA readily, generating a variety of DNA damages such as oxidized bases and strand breaks. The major mechanism that cells use to repair oxidized bases such as 8-hydroxyguanine, formamidopyrimidine and 5-hydroxyuracil is base excision repair (BER).[14] BER occurs in both the cell nucleus and in mitochondria.
References
- Li X, Fang P, Mai J, et al. (February 2013). "Targeting mitochondrial reactive oxygen species as novel therapy for inflammatory diseases and cancers". J Hematol Oncol. 6 (19): 19. doi:10.1186/1756-8722-6-19. PMC 3599349. PMID 23442817.
- Reichart, Gesine (October 30, 2018). "Mitochondrial complex IV mutation increases ROS production and reduces lifespan in aged mice". Acta Physiologica. 225: e13214. doi:10.1111/apha.13214. PMID 30376218.
- Li X, Fang P, et al. (March 2017). "Mitochondrial ROS, uncoupled from ATP synthesis, determine endothelial activation for both physiological recruitment of patrolling cells and pathological recruitment of inflammatory cells". Can J Physiol Pharmacol. 95 (3): 247–252. doi:10.1139/cjpp-2016-0515. PMC 5336492. PMID 27925481.
- Trewin, Adam J; Bahr, Laura L; Almast, Anmol; Berry, Brandon J; Wei, Alicia Y; Foster, Thomas H; Wojtovich, Andrew P (2019-03-19). "Mitochondrial ROS generated at the complex-II matrix or intermembrane space microdomain have distinct effects on redox signaling and stress sensitivity in C. elegans". Antioxidants & Redox Signaling. doi:10.1089/ars.2018.7681. ISSN 1523-0864.
- Trewin, Adam J.; Parker, Lewan; Shaw, Christopher S.; Hiam, Danielle S.; Garnham, Andrew; Levinger, Itamar; McConell, Glenn K.; Stepto, Nigel K. (November 2018). "Acute HIIE elicits similar changes in human skeletal muscle mitochondrial H 2 O 2 release, respiration, and cell signaling as endurance exercise even with less work". American Journal of Physiology. Regulatory, Integrative and Comparative Physiology. 315 (5): R1003–R1016. doi:10.1152/ajpregu.00096.2018. ISSN 0363-6119.
- Goncalves, Renata L. S.; Quinlan, Casey L.; Perevoshchikova, Irina V.; Hey-Mogensen, Martin; Brand, Martin D. (2015-01-02). "Sites of Superoxide and Hydrogen Peroxide Production by Muscle Mitochondria Assessed ex Vivo under Conditions Mimicking Rest and Exercise". Journal of Biological Chemistry. 290 (1): 209–227. doi:10.1074/jbc.M114.619072. ISSN 0021-9258. PMC 4281723. PMID 25389297.
- Li X, Fang P, Li Y, Kuo YM, Andrews AJ, Nanayakkara G, Johnson C, Fu H, Shan H, Du F, Hoffman NE, Yu D, Eguchi S, Madesh M, Koch WJ, Sun J, Jiang X, Wang H, Yang X (April 2016). "Mitochondrial Reactive Oxygen Species Mediate Lysophosphatidylcholine-Induced Endothelial Cell Activation". Arteriosclerosis, Thrombosis, and Vascular Biology. 36 (6): 1090–100. doi:10.1161/ATVBAHA.115.306964. PMC 4882253. PMID 27127201.
- West AP (April 2011). "TLR signalling augments macrophage bactericidal activity through mitochondrial ROS". Nature. 472 (7344): 476–480. doi:10.1038/nature09973. PMC 3460538. PMID 21525932.
- Finkel T (February 2012). "Signal transduction by mitochondrial oxidants". J Biol Chem. 287 (7): 4434–40. doi:10.1074/jbc.R111.271999. PMC 3281633. PMID 21832045.
- Cavounidis A, Mann EH (June 2020). "SARS-CoV-2 has a sweet tooth". Nature Reviews Immunology. doi:10.2139/ssrn.3606770. PMC 7291939. PMID 32533110.
- Velarde MC, Flynn JM, Day NU, Melov S, Campisi J (January 2012). "Mitochondrial oxidative stress caused by Sod2 deficiency promotes cellular senescence and aging phenotypes in the skin". Aging (Albany NY). 4 (1): 3–12. doi:10.18632/aging.100423. PMC 3292901. PMID 22278880.
- Velarde MC, Demaria M, Melov S, Campisi J (August 2015). "Pleiotropic age-dependent effects of mitochondrial dysfunction on epidermal stem cells". Proc. Natl. Acad. Sci. U.S.A. 112 (33): 10407–12. doi:10.1073/pnas.1505675112. PMC 4547253. PMID 26240345.
- Treiber N, Maity P, Singh K, Kohn M, Keist AF, Ferchiu F, Sante L, Frese S, Bloch W, Kreppel F, Kochanek S, Sindrilaru A, Iben S, Högel J, Ohnmacht M, Claes LE, Ignatius A, Chung JH, Lee MJ, Kamenisch Y, Berneburg M, Nikolaus T, Braunstein K, Sperfeld AD, Ludolph AC, Briviba K, Wlaschek M, Florin L, Angel P, Scharffetter-Kochanek K (April 2011). "Accelerated aging phenotype in mice with conditional deficiency for mitochondrial superoxide dismutase in the connective tissue". Aging Cell. 10 (2): 239–54. doi:10.1111/j.1474-9726.2010.00658.x. PMID 21108731.
- Maynard S, Schurman SH, Harboe C, de Souza-Pinto NC, Bohr VA (January 2009). "Base excision repair of oxidative DNA damage and association with cancer and aging". Carcinogenesis. 30 (1): 2–10. doi:10.1093/carcin/bgn250. PMC 2639036. PMID 18978338.