MDA5
MDA5 (melanoma differentiation-associated protein 5) is a RIG-I-like receptor dsRNA helicase enzyme that is encoded by the IFIH1 gene in humans.[5] MDA5 is part of the RIG-I-like receptor (RLR) family, which also includes RIG-I and LGP2, and functions as a pattern recognition receptor capable of detecting viruses. It is generally believed that MDA5 recognizes double stranded RNA (dsRNA) over 2000nts in length,[6] however it has been shown that whilst MDA5 can detect and bind to cytoplasmic dsRNA, it is also activated by a high molecular weight RNA complex composed of ssRNA and dsRNA.[7] For many viruses, effective MDA5-mediated antiviral responses are dependent on functionally active LGP2.[8] The signaling cascades in MDA5 is initiated via CARD domain.[9] Some observations made in cancer cells show that MDA5 also interacts with cellular RNA is able to induce an autoinflammatory response.[10]



Function
As a pattern recognition receptor
MDA5 is able to detect long dsRNA the genomic RNA of dsRNA viruses as well as replicative intermediates of both positive and negative sense RNA viruses.[11] MDA5 has also been shown to interact with a number of chemical modifications of RNA. The eukaryotic messenger RNA, for example, is often methylated at the 2’-O position of the first and second nucleotide behind the 5’ cap.[12] These structures are termed cap1 and cap2 respectively.[13] MDA5 is able to detect the absence of the 2'-O-methylation, bind to this type of RNA and initiate an immune response.[14]
Mechanism
Activated MDA5 interacts with the mitochondrial antiviral signalling proteins (MAVS) through its caspase activation and recruitment domains (CARDs) at the N-terminus.[15] The MAVS then work as a multiprotein complex to recruit the inhibitor of nuclear factor kappa-B kinase subunit epsilon (IKKε) along with the serine/threonine-protein kinase 1 (TBK1).[16] This causes the phosphorylation and the transport of the interferon regulatory factors 3 and 7 (IRF3 and IRF7) into the cell‘s nucleus. Once there, the regulatory factors induce the transcription of type I interferon genes IFN-β and IFN-α.[17]
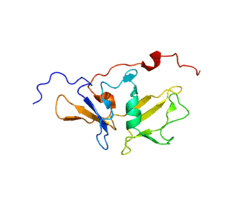
Structure
MDA5 is classified as an ATP-dependent DExD/H box RNA helicase. It comprises 2 CARD domains located at the N-terminus, a hinge region and the helicase domain which is made up of the domains RecA-like Hel1and Hel2.[18] Another hinge region connects the C-terminal domain (CTD) which is responsible for the recognition and the binding of RNA.[19] Apart from the positively charged groove recognizig the RNA, the CTD also contains a zinc binding domain.[20]
DEAD box proteins, characterized by the conserved motif Asp-Glu-Ala-Asp (DEAD), are putative RNA helicases. They are implicated in a number of cellular processes involving alteration of RNA secondary structure such as translation initiation, nuclear and mitochondrial splicing, and ribosome and spliceosome assembly. Based on their distribution patterns, some members of this family are believed to be involved in embryogenesis, spermatogenesis, and cellular growth and division. This gene encodes a DEAD box protein that is upregulated in response to treatment with beta-interferon (IFN-β) and a protein kinase C-activating compound, mezerein (MEZ). Irreversible reprogramming of melanomas can be achieved by treatment with both these agents; treatment with either agent alone only achieves reversible differentiation.[5]
Clinical significance
Mutations in IFIH1/MDA5 are associated to Singleton-Merten Syndrome[21] and to Aicardi–Goutières syndrome.
Some IFIH1 SNPs are associated with increased risk of type 1 diabetes.[22]
Antibodies against MDA5 are associated to amyopathic dermatomyositis with rapidly progressive interstitial lung disease.
References
- GRCh38: Ensembl release 89: ENSG00000115267 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000026896 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Entrez Gene: IFIH1 interferon induced with helicase C domain 1".
- Kato H, Takeuchi O, Mikamo-Satoh E, Hirai R, Kawai T, Matsushita K, et al. (July 2008). "Length-dependent recognition of double-stranded ribonucleic acids by retinoic acid-inducible gene-I and melanoma differentiation-associated gene 5". The Journal of Experimental Medicine. 205 (7): 1601–10. doi:10.1084/jem.20080091. PMC 2442638. PMID 18591409.
- Pichlmair A, Schulz O, Tan CP, Rehwinkel J, Kato H, Takeuchi O, et al. (October 2009). "Activation of MDA5 requires higher-order RNA structures generated during virus infection". Journal of Virology. 83 (20): 10761–9. doi:10.1128/JVI.00770-09. PMC 2753146. PMID 19656871.
- Satoh T, Kato H, Kumagai Y, Yoneyama M, Sato S, Matsushita K, et al. (January 2010). "LGP2 is a positive regulator of RIG-I- and MDA5-mediated antiviral responses". Proceedings of the National Academy of Sciences of the United States of America. 107 (4): 1512–7. Bibcode:2010PNAS..107.1512S. doi:10.1073/pnas.0912986107. PMC 2824407. PMID 20080593.
- Takeuchi O, Akira S (February 2008). "MDA5/RIG-I and virus recognition". Current Opinion in Immunology. 20 (1): 17–22. doi:10.1016/j.coi.2008.01.002. PMID 18272355.
- Dias Junior AG, Sampaio NG, Rehwinkel J (January 2019). "A Balancing Act: MDA5 in Antiviral Immunity and Autoinflammation". Trends in Microbiology. 27 (1): 75–85. doi:10.1016/j.tim.2018.08.007. PMC 6319154. PMID 30201512.
- Wu B, Peisley A, Richards C, Yao H, Zeng X, Lin C, et al. (January 2013). "Structural basis for dsRNA recognition, filament formation, and antiviral signal activation by MDA5". Cell. 152 (1–2): 276–89. doi:10.1016/j.cell.2012.11.048. PMID 23273991.
- Werner M, Purta E, Kaminska KH, Cymerman IA, Campbell DA, Mittra B, et al. (June 2011). "2'-O-ribose methylation of cap2 in human: function and evolution in a horizontally mobile family". Nucleic Acids Research. 39 (11): 4756–68. doi:10.1093/nar/gkr038. PMC 3113572. PMID 21310715.
- Byszewska M, Śmietański M, Purta E, Bujnicki JM (2014-12-02). "RNA methyltransferases involved in 5' cap biosynthesis". RNA Biology. 11 (12): 1597–607. doi:10.1080/15476286.2015.1004955. PMC 4615557. PMID 25626080.
- Züst R, Cervantes-Barragan L, Habjan M, Maier R, Neuman BW, Ziebuhr J, et al. (February 2011). "Ribose 2'-O-methylation provides a molecular signature for the distinction of self and non-self mRNA dependent on the RNA sensor Mda5". Nature Immunology. 12 (2): 137–43. doi:10.1038/ni.1979. PMC 3182538. PMID 21217758.
- Reikine S, Nguyen JB, Modis Y (2014). "Pattern Recognition and Signaling Mechanisms of RIG-I and MDA5". Frontiers in Immunology. 5: 342. doi:10.3389/fimmu.2014.00342. PMC 4107945. PMID 25101084.
- Fang R, Jiang Q, Zhou X, Wang C, Guan Y, Tao J, et al. (November 2017). "MAVS activates TBK1 and IKKε through TRAFs in NEMO dependent and independent manner". PLOS Pathogens. 13 (11): e1006720. doi:10.1371/journal.ppat.1006720. PMC 5699845. PMID 29125880.
- Brisse M, Ly H (2019). "Comparative Structure and Function Analysis of the RIG-I-Like Receptors: RIG-I and MDA5". Frontiers in Immunology. 10: 1586. doi:10.3389/fimmu.2019.01586. PMC 6652118. PMID 31379819.
- Rawling DC, Pyle AM (April 2014). "Parts, assembly and operation of the RIG-I family of motors". Current Opinion in Structural Biology. 25: 25–33. doi:10.1016/j.sbi.2013.11.011. PMC 4070197. PMID 24878341.
- Yoneyama M, Kikuchi M, Natsukawa T, Shinobu N, Imaizumi T, Miyagishi M, et al. (July 2004). "The RNA helicase RIG-I has an essential function in double-stranded RNA-induced innate antiviral responses". Nature Immunology. 5 (7): 730–7. doi:10.1038/ni1087. PMID 15208624.
- Cui S, Eisenächer K, Kirchhofer A, Brzózka K, Lammens A, Lammens K, et al. (February 2008). "The C-terminal regulatory domain is the RNA 5'-triphosphate sensor of RIG-I". Molecular Cell. 29 (2): 169–79. doi:10.1016/j.molcel.2007.10.032. PMID 18243112.
- Rutsch F, MacDougall M, Lu C, Buers I, Mamaeva O, Nitschke Y, et al. (February 2015). "A specific IFIH1 gain-of-function mutation causes Singleton-Merten syndrome". American Journal of Human Genetics. 96 (2): 275–82. doi:10.1016/j.ajhg.2014.12.014. PMC 4320263. PMID 25620204.
- Oram RA, Patel K, Hill A, Shields B, McDonald TJ, Jones A, et al. (March 2016). "A Type 1 Diabetes Genetic Risk Score Can Aid Discrimination Between Type 1 and Type 2 Diabetes in Young Adults". Diabetes Care. 39 (3): 337–44. doi:10.2337/dc15-1111. PMC 5642867. PMID 26577414.
Further reading
- Kang DC, Gopalkrishnan RV, Wu Q, Jankowsky E, Pyle AM, Fisher PB (January 2002). "mda-5: An interferon-inducible putative RNA helicase with double-stranded RNA-dependent ATPase activity and melanoma growth-suppressive properties". Proceedings of the National Academy of Sciences of the United States of America. 99 (2): 637–42. Bibcode:2002PNAS...99..637K. doi:10.1073/pnas.022637199. PMC 117358. PMID 11805321.
- Kovacsovics M, Martinon F, Micheau O, Bodmer JL, Hofmann K, Tschopp J (May 2002). "Overexpression of Helicard, a CARD-containing helicase cleaved during apoptosis, accelerates DNA degradation". Current Biology. 12 (10): 838–43. doi:10.1016/S0960-9822(02)00842-4. PMID 12015121.
- Cocude C, Truong MJ, Billaut-Mulot O, Delsart V, Darcissac E, Capron A, et al. (December 2003). "A novel cellular RNA helicase, RH116, differentially regulates cell growth, programmed cell death and human immunodeficiency virus type 1 replication". The Journal of General Virology. 84 (Pt 12): 3215–3225. doi:10.1099/vir.0.19300-0. PMID 14645903.
- Kang DC, Gopalkrishnan RV, Lin L, Randolph A, Valerie K, Pestka S, Fisher PB (March 2004). "Expression analysis and genomic characterization of human melanoma differentiation associated gene-5, mda-5: a novel type I interferon-responsive apoptosis-inducing gene". Oncogene. 23 (9): 1789–800. doi:10.1038/sj.onc.1207300. PMID 14676839.
- Andrejeva J, Childs KS, Young DF, Carlos TS, Stock N, Goodbourn S, Randall RE (December 2004). "The V proteins of paramyxoviruses bind the IFN-inducible RNA helicase, mda-5, and inhibit its activation of the IFN-beta promoter". Proceedings of the National Academy of Sciences of the United States of America. 101 (49): 17264–9. Bibcode:2004PNAS..10117264A. doi:10.1073/pnas.0407639101. PMC 535396. PMID 15563593.
- Yoneyama M, Kikuchi M, Matsumoto K, Imaizumi T, Miyagishi M, Taira K, et al. (September 2005). "Shared and unique functions of the DExD/H-box helicases RIG-I, MDA5, and LGP2 in antiviral innate immunity". Journal of Immunology. 175 (5): 2851–8. doi:10.4049/jimmunol.175.5.2851. PMID 16116171.
- Kawai T, Takahashi K, Sato S, Coban C, Kumar H, Kato H, et al. (October 2005). "IPS-1, an adaptor triggering RIG-I- and Mda5-mediated type I interferon induction". Nature Immunology. 6 (10): 981–8. doi:10.1038/ni1243. PMID 16127453.
- Meylan E, Curran J, Hofmann K, Moradpour D, Binder M, Bartenschlager R, Tschopp J (October 2005). "Cardif is an adaptor protein in the RIG-I antiviral pathway and is targeted by hepatitis C virus". Nature. 437 (7062): 1167–72. Bibcode:2005Natur.437.1167M. doi:10.1038/nature04193. PMID 16177806.
- Lin L, Su Z, Lebedeva IV, Gupta P, Boukerche H, Rai T, et al. (November 2006). "Activation of Ras/Raf protects cells from melanoma differentiation-associated gene-5-induced apoptosis". Cell Death and Differentiation. 13 (11): 1982–93. doi:10.1038/sj.cdd.4401899. PMID 16575407.
- Smyth DJ, Cooper JD, Bailey R, Field S, Burren O, Smink LJ, et al. (June 2006). "A genome-wide association study of nonsynonymous SNPs identifies a type 1 diabetes locus in the interferon-induced helicase (IFIH1) region". Nature Genetics. 38 (6): 617–9. doi:10.1038/ng1800. PMID 16699517.
- Berghäll H, Sirén J, Sarkar D, Julkunen I, Fisher PB, Vainionpää R, Matikainen S (July 2006). "The interferon-inducible RNA helicase, mda-5, is involved in measles virus-induced expression of antiviral cytokines". Microbes and Infection. 8 (8): 2138–44. doi:10.1016/j.micinf.2006.04.005. PMID 16782388.
- Sirén J, Imaizumi T, Sarkar D, Pietilä T, Noah DL, Lin R, et al. (July 2006). "Retinoic acid inducible gene-I and mda-5 are involved in influenza A virus-induced expression of antiviral cytokines". Microbes and Infection. 8 (8): 2013–20. doi:10.1016/j.micinf.2006.02.028. PMID 16797201.
- Barral PM, Morrison JM, Drahos J, Gupta P, Sarkar D, Fisher PB, Racaniello VR (April 2007). "MDA-5 is cleaved in poliovirus-infected cells". Journal of Virology. 81 (8): 3677–84. doi:10.1128/JVI.01360-06. PMC 1866155. PMID 17267501.
- Marinou I, Montgomery DS, Dickson MC, Binks MH, Moore DJ, Bax DE, Wilson AG (2007). "The interferon induced with helicase domain 1 A946T polymorphism is not associated with rheumatoid arthritis". Arthritis Research & Therapy. 9 (2): R40. doi:10.1186/ar2179. PMC 1906818. PMID 17442111.
- Sutherland A, Davies J, Owen CJ, Vaikkakara S, Walker C, Cheetham TD, et al. (August 2007). "Genomic polymorphism at the interferon-induced helicase (IFIH1) locus contributes to Graves' disease susceptibility". The Journal of Clinical Endocrinology and Metabolism. 92 (8): 3338–41. doi:10.1210/jc.2007-0173. PMC 6952273. PMID 17535987.
- Matsukura S, Kokubu F, Kurokawa M, Kawaguchi M, Ieki K, Kuga H, et al. (2007). "Role of RIG-I, MDA-5, and PKR on the expression of inflammatory chemokines induced by synthetic dsRNA in airway epithelial cells". International Archives of Allergy and Immunology. 143 Suppl 1: 80–3. doi:10.1159/000101411. PMID 17541283.