Gap-43 protein
Growth Associated Protein 43 (GAP43) is a protein encoded by the GAP43 gene[5] in humans.

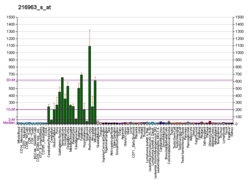
GAP43 is called a "growth" or "plasticity" protein because it is expressed at high levels in neuronal growth cones during development[6] and axonal regeneration, and it is phosphorylated after long-term potentiation and after learning.
GAP43 is a crucial component of the axon and presynaptic terminal. Its null mutation leads to death within days after birth, due to axon pathfinding defects.[7]
Synonyms
GAP43 is also referred to as:
- protein F1
- neuromodulin
- neural phosphoprotein B-50
- axonal membrane protein GAP-43
- calmodulin-binding protein P-57
- nerve growth-related peptide GAP43
- neuron growth-associated protein 43
Function
GAP43, is a nervous tissue-specific cytoplasmic protein that can be attached to the membrane via a dual palmitoylation sequence on cysteines 3 and 4. This sequence targets GAP43 to lipid rafts. It is a major protein kinase C (PKC) substrate and is considered to play a key role in neurite formation, regeneration, and plasticity.[8][9] The role of GAP-43 in CNS development is not limited to effects on axons: It is also a component of the centrosome, and differentiating neurons that do not express GAP-43 show mislocalization of the centrosome and mitotic spindles, particularly in neurogenic cell divisions. As a consequence, in the cerebellum, the neuronal precursor pool fails to expand normally and the cerebellum is significantly smaller.[10]
Several different laboratories studying the same protein, now called GAP43, initially used different names. It was designated F1, then B-50, then GAP43, pp46, and finally neuromodulin, each name reflecting a different function of the same molecule.[11] F1 was localized to synapses, and was increased in its phosphorylation one day after learning. However, F1 was not cAMP kinase dependent. B-50 was regulated by the pituitary peptide ACTH and was associated with grooming behavior. In the case of GAP-43, it was designated as a growth-associated protein because its synthesis was upregulated during axonal regeneration. Pp46 was concentrated in neuronal growth cones and was thus postulated to play an important role in brain development. In the case of neuromodulin, it was shown to bind calmodulin avidly.
GAP43, the consensus choice for its designation,[11] is a nervous system-specific protein that is attached to the membrane via a dual palmitoylation sequence on cysteines 3 and 4, though it can exist in the non-bound form in the cytoplasm. This dual sequence enables the association of phosphatidylinositol-4,5-bisphosphate [PI(4,5)P2] or PIP2, with actin, facilitating the latter’s polymerization thereby regulating neuronal structure. This can occur within a lipid raft so as to compartmentalize and localize motility of filopodia in growth cones in developing brains, and could also remodel presynaptic terminals in adults in an activity-dependent manner. GAP-43 is also a protein kinase C (PKC) substrate. Phosphorylation of serine-41 on GAP-43 by PKC regulates neurite formation, regeneration, and synaptic plasticity.[8]
Because of the association and potential binding of GAP43 with a number of different molecules, including PKC, PIP2, actin, calmodulin, spectrin, palmitate, synaptophysin, amyloid and tau protein, it may be useful to think of GAP43 as an adaptor protein situated within the terminal in a supramolecular complex regulating presynaptic terminal functions, particularly bidirectional communication with the postsynaptic process. Its important role in memory and information storage is executed through its cell biological mechanisms of phosphorylation, palmitoylation, protein-protein interaction and structural remodeling via actin polymerization.
Clinical significance
Humans with a deletion in one allele of the GAP43 gene fail to form telencephalic commissures and are intellectually disabled.[12][13]
Model organisms
| Characteristic | Phenotype |
|---|---|
| Homozygote viability | Abnormal |
| Recessive lethal study | Normal |
| Fertility | Normal |
| Body weight | Normal |
| Anxiety | Normal |
| Neurological assessment | Normal |
| Grip strength | Normal |
| Hot plate | Normal |
| Dysmorphology | Normal |
| Indirect calorimetry | Normal |
| Glucose tolerance test | Normal |
| Auditory brainstem response | Normal |
| DEXA | Normal |
| Radiography | Normal |
| Body temperature | Normal |
| Eye morphology | Normal |
| Clinical chemistry | Normal |
| Plasma immunoglobulins | Abnormal |
| Haematology | Normal |
| Micronucleus test | Normal |
| Heart weight | Normal |
| Brain histopathology | Normal |
| Eye Histopathology | Normal |
| Salmonella infection | Normal[14] |
| Citrobacter infection | Normal[15] |
| All tests and analysis from[16][17] |
Model organisms have been used in the study of GAP43 function. A conditional knockout mouse line, called Gap43tm1a(EUCOMM)Wtsi[18][19] was generated as part of the International Knockout Mouse Consortium program—a high-throughput mutagenesis project to generate and distribute animal models of disease to interested scientists.[20][21][22] Male and female animals underwent a standardized phenotypic screen to determine the effects of deletion.[16][23] Twenty five tests were carried out on mutant mice and two significant abnormalities were observed. No homozygous mutant mice survived until weaning. The remaining tests were carried out on heterozygous mutant adult mice and increased IgG1 levels were observed in these animals.[16]
Studies on another homozygous GAP43 knockout mouse line found it to be lethal days after birth because it plays a critical role in the development of the mammalian CNS.[24] Telencephalic commissures fail to form,[25] thalamocortical afferents are mistargeted, especially in somatosensory, particularly barrel, cortex.[24] GAP43 is not only important for axon targeting during development but it has been shown to be important also for the maintenance of the structure and dynamics of axonal fibres and of their synaptic terminals in wild-type rodents both during normal conditions and during lesion-induced axonal sprouting.[26][27][28] The cerebellum is also affected.[29] GAP43 is also haploinsufficient for the cortical phenotypes and the severity of the axon targeting phenotype is directly related to the extent to which the affected axons are phosphorylated by PKC, suggesting that axons require a functional threshold of phosphorylated GAP43 for targeting to occur normally.[25] Moreover, elevation above this threshold in GAP43 mice can enhance learning and also facilitate a physiological model of learning, long-term potentiation (LTP).[30] However, further enrichment beyond a certain level can be devastating to cognitive functions.
References
- GRCh38: Ensembl release 89: ENSG00000172020 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000047261 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Kosik KS, Orecchio LD, Bruns GA, Benowitz LI, MacDonald GP, Cox DR, Neve RL (Apr 1988). "Human GAP-43: its deduced amino acid sequence and chromosomal localization in mouse and human". Neuron. 1 (2): 127–32. doi:10.1016/0896-6273(88)90196-1. PMID 3272162.
- Referenced within : Rosskothen-Kuhl N, Illing RB (2014). "Gap43 Transcription Modulation in the Adult Brain Depends on Sensory Activity and Synaptic Cooperation". PLoS ONE. 9 (3): e92624. doi:10.1371/journal.pone.0092624. PMC 3960265. PMID 24647228.
- "Entrez Gene: GAP43 growth associated protein 43".
- Benowitz LI, Routtenberg A (Feb 1997). "GAP-43: an intrinsic determinant of neuronal development and plasticity". Trends in Neurosciences. 20 (2): 84–91. doi:10.1016/S0166-2236(96)10072-2. PMID 9023877.
- Aarts LH, Schotman P, Verhaagen J, Schrama LH, Gispen WH (1998). The role of the neural growth associated protein B-50/GAP-43 in morphogenesis. Advances in Experimental Medicine and Biology. 446. pp. 85–106. doi:10.1007/978-1-4615-4869-0_6. ISBN 978-1-4613-7209-7. PMID 10079839.
- Mishra R, ManiS (2008). "GAP-43 is key to mitotic spindle control and centrosome-based polarization in neurons". Cell Cycle. 7 (3): 348–357. doi:10.4161/cc.7.3.5235. PMID 18235238.
- Benowitz LI, Routtenberg A (1987). "A membrane phosphoprotein associated with neural development, axonal regeneration, phospholipid metabolism, and synaptic plasticity". Trends in Neurosciences. 10 (12): 527–532. doi:10.1016/0166-2236(87)90135-4.
- Genuardi M, Calvieri F, Tozzi C, Coslovi R, Neri G (Oct 1994). "A new case of interstitial deletion of chromosome 3q, del(3q)(q13.12q21.3), with agenesis of the corpus callosum". Clinical Dysmorphology. 3 (4): 292–6. doi:10.1097/00019605-199410000-00003. PMID 7894733.
- Mackie Ogilvie C, Rooney SC, Hodgson SV, Berry AC (Mar 1998). "Deletion of chromosome 3q proximal region gives rise to a variable phenotype". Clinical Genetics. 53 (3): 220–2. doi:10.1111/j.1399-0004.1998.tb02681.x. PMID 9630079.
- "Salmonella infection data for Gap43". Wellcome Trust Sanger Institute.
- "Citrobacter infection data for Gap43". Wellcome Trust Sanger Institute.
- Gerdin AK (2010). "The Sanger Mouse Genetics Programme: High throughput characterisation of knockout mice". Acta Ophthalmologica. 88: 925–7. doi:10.1111/j.1755-3768.2010.4142.x.
- Mouse Resources Portal, Wellcome Trust Sanger Institute.
- "International Knockout Mouse Consortium".
- "Mouse Genome Informatics".
- Skarnes WC, Rosen B, West AP, Koutsourakis M, Bushell W, Iyer V, Mujica AO, Thomas M, Harrow J, Cox T, Jackson D, Severin J, Biggs P, Fu J, Nefedov M, de Jong PJ, Stewart AF, Bradley A (Jun 2011). "A conditional knockout resource for the genome-wide study of mouse gene function". Nature. 474 (7351): 337–42. doi:10.1038/nature10163. PMC 3572410. PMID 21677750.
- Dolgin E (Jun 2011). "Mouse library set to be knockout". Nature. 474 (7351): 262–3. doi:10.1038/474262a. PMID 21677718.
- Collins FS, Rossant J, Wurst W (Jan 2007). "A mouse for all reasons". Cell. 128 (1): 9–13. doi:10.1016/j.cell.2006.12.018. PMID 17218247.
- van der Weyden L, White JK, Adams DJ, Logan DW (2011). "The mouse genetics toolkit: revealing function and mechanism". Genome Biology. 12 (6): 224. doi:10.1186/gb-2011-12-6-224. PMC 3218837. PMID 21722353.
- Strittmatter SM, Fankhauser C, Huang PL, Mashimo H, Fishman MC (Feb 1995). "Neuronal pathfinding is abnormal in mice lacking the neuronal growth cone protein GAP-43". Cell. 80 (3): 445–52. doi:10.1016/0092-8674(95)90495-6. PMID 7859286.
- Shen Y, Mani S, Donovan SL, Schwob JE, Meiri KF (Jan 2002). "Growth-associated protein-43 is required for commissural axon guidance in the developing vertebrate nervous system". The Journal of Neuroscience. 22 (1): 239–47. doi:10.1523/JNEUROSCI.22-01-00239.2002. PMC 6757587. PMID 11756507.
- Grasselli G, Mandolesi G, Strata P, Cesare P (June 2011). "Impaired sprouting and axonal atrophy in cerebellar climbing fibres following in vivo silencing of the growth-associated protein GAP-43". PLOS ONE. 6 (6): e20791. doi:10.1371/journal.pone.0020791. PMC 3112224. PMID 21695168.
- Grasselli G, Strata P (February 2013). "Structural plasticity of climbing fibers and the growth-associated protein GAP-43". Frontiers in Neural Circuits. 7 (25): 25. doi:10.3389/fncir.2013.00025. PMC 3578352. PMID 23441024.
- Allegra Mascaro AL, Cesare P, Sacconi L, Grasselli G, Mandolesi G, Maco B, Knott GW, Huang L, De Paola V, Strata P, Pavone FS (Jun 2013). "In vivo single branch axotomy induces GAP-43-dependent sprouting and synaptic remodeling in cerebellar cortex". Proceedings of the National Academy of Sciences of the United States of America. 110 (26): 10824–9. doi:10.1073/pnas.1219256110. PMC 3696745. PMID 23754371.
- Shen Y, Mishra R, Mani S, Meiri KF (2008). "Both cell-autonomous and cell non-autonomous functions of GAP-43 are required for normal patterning of the cerebellum in vivo". Cerebellum. 7 (3): 451–66. doi:10.1007/s12311-008-0049-5. PMC 4164963. PMID 18777197.
- Routtenberg A, Cantallops I, Zaffuto S, Serrano P, Namgung U (Jun 2000). "Enhanced learning after genetic overexpression of a brain growth protein". Proceedings of the National Academy of Sciences of the United States of America. 97 (13): 7657–62. doi:10.1073/pnas.97.13.7657. PMC 16601. PMID 10861025.
Further reading
- Fantini F, Johansson O (Dec 1992). "Expression of growth-associated protein 43 and nerve growth factor receptor in human skin: a comparative immunohistochemical investigation". The Journal of Investigative Dermatology. 99 (6): 734–42. doi:10.1111/1523-1747.ep12614465. PMID 1281863.
- Mercken M, Lübke U, Vandermeeren M, Gheuens J, Oestreicher AB (Apr 1992). "Immunocytochemical detection of the growth-associated protein B-50 by newly characterized monoclonal antibodies in human brain and muscle". Journal of Neurobiology. 23 (3): 309–21. doi:10.1002/neu.480230310. PMID 1385623.
- Spencer SA, Schuh SM, Liu WS, Willard MB (May 1992). "GAP-43, a protein associated with axon growth, is phosphorylated at three sites in cultured neurons and rat brain". The Journal of Biological Chemistry. 267 (13): 9059–64. PMID 1533624.
- Apel ED, Litchfield DW, Clark RH, Krebs EG, Storm DR (Jun 1991). "Phosphorylation of neuromodulin (GAP-43) by casein kinase II. Identification of phosphorylation sites and regulation by calmodulin". The Journal of Biological Chemistry. 266 (16): 10544–51. PMID 1828073.
- Apel ED, Byford MF, Au D, Walsh KA, Storm DR (Mar 1990). "Identification of the protein kinase C phosphorylation site in neuromodulin". Biochemistry. 29 (9): 2330–5. doi:10.1021/bi00461a017. PMID 2140056.
- Kosik KS, Orecchio LD, Bruns GA, Benowitz LI, MacDonald GP, Cox DR, Neve RL (Apr 1988). "Human GAP-43: its deduced amino acid sequence and chromosomal localization in mouse and human". Neuron. 1 (2): 127–32. doi:10.1016/0896-6273(88)90196-1. PMID 3272162.
- Ng SC, de la Monte SM, Conboy GL, Karns LR, Fishman MC (Apr 1988). "Cloning of human GAP-43: growth association and ischemic resurgence". Neuron. 1 (2): 133–9. doi:10.1016/0896-6273(88)90197-3. PMID 3272163.
- Nielander HB, De Groen PC, Eggen BJ, Schrama LH, Gispen WH, Schotman P (Sep 1993). "Structure of the human gene for the neural phosphoprotein B-50 (GAP-43)". Brain Research. Molecular Brain Research. 19 (4): 293–302. doi:10.1016/0169-328X(93)90128-C. hdl:1874/4067. PMID 8231732.
- Oehrlein SA, Parker PJ, Herget T (Jul 1996). "Phosphorylation of GAP-43 (growth-associated protein of 43 kDa) by conventional, novel and atypical isotypes of the protein kinase C gene family: differences between oligopeptide and polypeptide phosphorylation". The Biochemical Journal. 317. 317 (1): 219–24. doi:10.1042/bj3170219. PMC 1217466. PMID 8694767.
- Kanazir S, Ruzdijic S, Vukosavic S, Ivkovic S, Milosevic A, Zecevic N, Rakic L (May 1996). "GAP-43 mRNA expression in early development of human nervous system". Brain Research. Molecular Brain Research. 38 (1): 145–55. doi:10.1016/0169-328X(96)00008-3. PMID 8737678.
- de Groen PC, Eggen BJ, Gispen WH, Schotman P, Schrama LH (1996). "Cloning and promoter analysis of the human B-50/GAP-43 gene". Journal of Molecular Neuroscience. 6 (2): 109–19. doi:10.1007/BF02736770. PMID 8746449.
- Chao S, Benowitz LI, Krainc D, Irwin N (Sep 1996). "Use of a two-hybrid system to investigate molecular interactions of GAP-43". Brain Research. Molecular Brain Research. 40 (2): 195–202. doi:10.1016/0169-328X(96)00049-6. PMID 8872303.
- Gamby C, Waage MC, Allen RG, Baizer L (Oct 1996). "Analysis of the role of calmodulin binding and sequestration in neuromodulin (GAP-43) function". The Journal of Biological Chemistry. 271 (43): 26698–705. doi:10.1074/jbc.271.43.26698. PMID 8900147.
- Heuss D, Schlötzer-Schrehardt U (Jun 1998). "Subcellular localization of phosphoprotein B-50 in regenerating muscle. An immuno-electron microscopic study". Neurological Research. 20 (4): 360–4. PMID 9618702.
- Neve RL, Coopersmith R, McPhie DL, Santeufemio C, Pratt KG, Murphy CJ, Lynn SD (Oct 1998). "The neuronal growth-associated protein GAP-43 interacts with rabaptin-5 and participates in endocytosis". The Journal of Neuroscience. 18 (19): 7757–67. doi:10.1523/JNEUROSCI.18-19-07757.1998. PMC 6793001. PMID 9742146.
- Arni S, Keilbaugh SA, Ostermeyer AG, Brown DA (Oct 1998). "Association of GAP-43 with detergent-resistant membranes requires two palmitoylated cysteine residues". The Journal of Biological Chemistry. 273 (43): 28478–85. doi:10.1074/jbc.273.43.28478. PMID 9774477.
- Eastwood SL, Harrison PJ (Sep 1998). "Hippocampal and cortical growth-associated protein-43 messenger RNA in schizophrenia". Neuroscience. 86 (2): 437–48. doi:10.1016/S0306-4522(98)00040-2. PMID 9881859.
- Cargill M, Altshuler D, Ireland J, Sklar P, Ardlie K, Patil N, Shaw N, Lane CR, Lim EP, Kalyanaraman N, Nemesh J, Ziaugra L, Friedland L, Rolfe A, Warrington J, Lipshutz R, Daley GQ, Lander ES (Jul 1999). "Characterization of single-nucleotide polymorphisms in coding regions of human genes". Nature Genetics. 22 (3): 231–8. doi:10.1038/10290. PMID 10391209.
- Riederer BM, Routtenberg A (Aug 1999). "Can GAP-43 interact with brain spectrin?". Brain Research. Molecular Brain Research. 71 (2): 345–8. doi:10.1016/S0169-328X(99)00179-5. PMID 10521589.
- Vento P, Soinila S (Nov 1999). "Quantitative comparison of growth-associated protein GAP-43, neuron-specific enolase, and protein gene product 9.5 as neuronal markers in mature human intestine". The Journal of Histochemistry and Cytochemistry. 47 (11): 1405–16. doi:10.1177/002215549904701107. PMID 10544214.
External links
- Gap-43+protein at the US National Library of Medicine Medical Subject Headings (MeSH)
- ihop-net, Growth associated protein 43
- NCBI