Fibril
Fibrils (from the Latin fibra[1]) are structural biological materials found in nearly all living organisms. Not to be confused with fibers or filaments, fibrils tend to have diameters ranging from 10-100 nanometers (whereas fibers are micro to milli-scale structures and filaments have diameters approximately 10-50 nanometers in size). Fibrils are not usually found alone but rather are parts of greater hierarchical structures commonly found in biological systems. Due to the prevalence of fibrils in biological systems, their study is of great importance in the fields of microbiology, biomechanics, and materials science.
Structure and mechanics
Fibrils are composed of linear biopolymers, and are characterized by rod-like structures with high length-to-diameter ratios. Oftentimes, they spontaneously arrange into helical structures. In biomechanics problems, fibrils can be characterized as classical beams with a roughly circular cross-sectional area on the nanometer scale. As such, simple beam bending equations can be applied to calculate flexural strength of fibrils under ultra-low loading conditions. Like most biopolymers, stress-strain relationships of fibrils tend to show a characteristic toe-heel region before a linear, elastic region.[2] Unlike biopolymers, fibrils do not behave like homogeneous materials, as yield strength has been shown to vary with volume, indicating structural dependencies.[3]
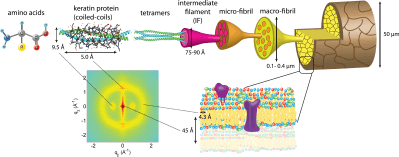
Differences in structure between fibrils of different origin is typically determined by x-ray diffraction.[2] A scanning electron microscope (SEM) can be used to observe specific details on larger fibril species such as the characteristic 67 nm bands in collagen, but often is not fine enough to determine the full structure.
Hydration has been shown to produce a noticeable effect in the mechanical properties of fibrillar materials. The presence of water has been shown to decrease the stiffness of collagen fibrils, as well as increase their rate of stress relaxation and strength.[4] From a biological standpoint, water content acts as a toughening mechanism for fibril structures, allowing for higher energy absorption and greater straining capabilities.
Fibrillogenesis
Fibrillogenesis is the expansion of fine fibrils which is common in collagen fibers of connective tissue. The definite mechanisms of fibrillogenesis are still unknown, although many hypotheses resulting from basic research help discover many possible mechanisms. In early experiments, collagen I could be distilled from tissues and recombined into fibrils with controlling the solutions. Later studies help understand the composition and structure of binding sites on the collagen monomers. Collagen is synthesized as a soluble precursor, procollagen, which supports collagen self-assembly. Since collagen fibrils have almost 50 binding components in vivo, the definite requirement to generate fibrillogenesis in vivo is still cryptic.[5]
With acidic or saline solution, collagen can be extracted from tissues and rearrange into fibril by changing temperature or pH value.[6] Experiments discovered attractive force between collagen monomers which helps the rearrangement.[7] Collagen serves as a precursor, procollagen, in synthesizing reaction, which identifies self-polymerization of collagen.
Natural processes
There are over 30 collagens in nature that are similar in chemical composition but differ in terms of crystal structure. By far, collagen I and II are the most abundant. They initiatively form fibrils in vitro, while fibronectin, fibronectin-binding, collagen-binding integrins and collagen V are essential for collagen I forming and collagen XI for collagen II forming. Therefore, cellular mechanisms play key role in the protein self-assembly process.
In animals
Collagen
.png)
Collagen is the major structural protein outside cells in many connective tissues of animals.[8] As the primary component of connective tissue, it has the largest amount among protein in mammals, occupying 25% to 35% of all protein content in the body.
The fibrils in collagen are packed in a crimp structure. The stress/strain curve of collagen, such as tendon, can be subdivided into several regions. The region of small strains,"toe"region, corresponds to the removal of a macroscopic crimp, uncrimping, in the collagen fibrils, visible in light microscope. At larger strains, "heel" and "linear" region, there's no further structural change visible.
Tropocollagen is the molecular component fiber, consisting of three left handed polypeptide chains (red, green, blue) coiled around each other, forming a right-handed triple helix.
Actin and myosin
Muscles contract and stretch via the steerable sliding/grasping of the myosin interacting with actin fibers. Actin consists of two polypeptides in a helix and myosin has a small heart-shaped structure, cross-bridge. The bind and unbind processes of cross-bridge attaching on actin filament help relative movement of these collagens and hence the whole muscle.
Elastin and keratin
Elastin is a fibrous protein common in various soft tissues, like skin, blood vessels and lung tissue. Each monomer connects with each other, forming a 3D network, with ability to endure over 200% strain before deformation.[9]
Keratin is a structural protein mainly found in hair, nails, hooves, horns, quills.[10] Basically keratin is formed by polypeptide chains, which coil into α-helices with sulfur cross-links or bond into β-sheets linked by hydrogen bonding. β-keratin, which is tougher than α-conformation, is more common in birds and reptiles.
Resilin and spider silk
Resilin is an elastomeric insect protein, consisting of both α-helices and β-sheets structure.[11] It is one of the most resilient protein in nature. It has a low stiffness ~0.6MPa but a high energy restoring percentage ~98%, and efficiently helps flying insects to flap wings or fleas to jump.
Spider silk fibril is composed of stiff crystallized β-sheets structure, responsible for strength, and amorphous matrix surrounding, improving toughness and elongation ability.[12] It has exceptionally high tensile strength and ductility, with respectively low density, compared to other natural fibril. Its feature varies from different kinds of spider for different utility.
In plants
Cellulose

The primary cell wall derives its notable tensile strength from cellulose molecules, or long-chains of glucose residues stabilized by hydrogen bonding.[13] Cellulose chains are observed to align in overlapping parallel arrays, with the similar polarity forming a cellulose microfibril. In plants, these cellulose microfibrils arrange themselves into layers, formally known as lamellae, and are stabilized in the cell wall by surface, long cross-linking glycan molecules. Glycan molecules increase the complexity of the potential networks plant-based cellulose can configure itself into. Coextensive in the primary cell wall to both cellulose microfibrils and complementary glycan networks, is pectin which is a polysaccharide that contains many negatively charged galacturonic acid units.[13] Additionally, cellulose microfibrils also contribute to the shape of the plant via controlled-cell expansion. The stereoscopic arrangement of microfibrils in the cell wall create systems of turgor pressure which ultimately leads to cellular growth and expansion. Cellulose microfibrils are unique matrix macromolecules, in that they are assembled by cellulose synthase enzymes located on the extracellular surface of the plasma membrane.[13] It is believed that the plant can “anticipate their future morphology by controlling the orientation of microfibrils” by a mechanism where cellulose microfibrils are arranged atop a cortical array of microtubules.
Amylose
Stirring of a given sample of amylose is said to form fibrillar crystals which are said to precipitate out of the mother liquor. These long fibrils can be imaged using electron microscopy revealing transverse striations resembling a shish-kebab. Amylose fibrils are categorized with having one of two morphologies: ones with small rodlike fibrils and others with lath-shaped crystals.
Wood
The fibrillar structure of wood is said to play a significant role in both the mechanical stability and ability of wood to possess channels to transport minerals and water. Sprucewood (Picea abies), among others, are reported to possess cellulose fibrils with a normalized diameter of 2.5 nm. There is also a reported link between the age of the wood and the spiral angle of the fibrils with respect to the longitudinal direction. Earlywood is said to have a consistent 4.6 ± 0.6° rest angle, whereas latewood is said to have a transition region from 4.6° to 19.8 ± 0.7°.[14] In latewood, the two spiral angle regions of cellulose fibrils are not continuous, meaning that there are two independent tracheid structures in “older” trees meeting different mechanical requirements. Moreover, longitudinally oriented fibrils improve tensile strength, whereas the addition of 20° tilted fibrils, exclusive to latewood tracheids, provides stability against compression.[14]
Biomimicry and fibrils
Self-cleaning properties
In order to mimic the strong adhesion, easy detachment, and self-cleaning properties of a gecko toe pad, a fibrillar-based adhesive can be created. These performance features stem from the underlying hierarchical structure which consists of a million microfibrils called setae which further consists of billions of nano-sized branches called spatulae.
Mimicking this phenomenon involves four distinct design steps:[15]
- Making a vertically aligned micro-/nano- fibrillar arrays
- Creating various tip shapes
- Including anisotropic geometry
- Building hierarchy.
A mature bone matrix
In order to mimic a mature bone matrix, self-assembled fibrils can be used to align a given mineral matrix. This is accomplished using a self-assembling molecule with a hydrophobic alkyl tail and a hydrophilic oligopeptide head. These molecules form micellar structures in situ, and disulfide bridges at low pH, leading to the formation and crystallization of 200 kDa polymeric nanofibrils. The mineral matrix ultimately interacts with the synthetic fibril via a phosphoserine residue which results in mineral nucleation and growth.
References
- "Online Etymology Dictionary". www.etymonline.com. Retrieved 2017-03-31.
- Fratzl, Peter (1998). "Fibrillar Structure and Mechanical Properties of Collagen". Journal of Structural Biology. 122 (1–2): 119–122. doi:10.1006/jsbi.1998.3966. PMID 9724612.
- Shen, Zhilei L.; Dodge, Mohammad Reza; Kahn, Harold; Ballarini, Roberto; Eppell, Steven J. (2008-10-15). "Stress-Strain Experiments on Individual Collagen Fibrils". Biophysical Journal. 95 (8): 3956–3963. Bibcode:2008BpJ....95.3956S. doi:10.1529/biophysj.107.124602. ISSN 0006-3495. PMC 2553131. PMID 18641067.
- Chimich, D. (1992). "Water content alters viscoelastic behaviour of the normal adolescent rabbit medial collateral ligament". Journal of Biomechanics. 25 (8): 831–837. doi:10.1016/0021-9290(92)90223-N.
- Kadler, Karl E; Hill, Adele; Canty-Laird, Elizabeth G (2017-05-02). "Collagen fibrillogenesis: fibronectin, integrins, and minor collagens as organizers and nucleators". Current Opinion in Cell Biology. 20 (5–24): 495–501. doi:10.1016/j.ceb.2008.06.008. ISSN 0955-0674. PMC 2577133. PMID 18640274.
- Gross, J.; Kirk, D. (1958-08-01). "The heat precipitation of collagen from neutral salt solutions: some rate-regulating factors". The Journal of Biological Chemistry. 233 (2): 355–360. ISSN 0021-9258. PMID 13563501.
- Prockop, D. J.; Fertala, A. (1998-06-19). "Inhibition of the self-assembly of collagen I into fibrils with synthetic peptides. Demonstration that assembly is driven by specific binding sites on the monomers". The Journal of Biological Chemistry. 273 (25): 15598–15604. doi:10.1074/jbc.273.25.15598. ISSN 0021-9258. PMID 9624151.
- Kadler, Karl E; Hill, Adele; Canty-Laird, Elizabeth G (2017-03-31). "Collagen fibrillogenesis: fibronectin, integrins, and minor collagens as organizers and nucleators". Current Opinion in Cell Biology. 20 (5–24): 495–501. doi:10.1016/j.ceb.2008.06.008. ISSN 0955-0674. PMC 2577133. PMID 18640274.
- Gosilin, J.M.; Aaron, B.B (1981). "Elastin as a Random-network elastomer, a mechanical and optical analysis of single elastin fibers". Biopolymers. 20 (6): 1247–1260. doi:10.1002/bip.1981.360200611.
- Meyers, M. A. (2014). Biology in Materials Science. Cambridge, UK: Cambridge University Press.
- Weis-Fough, T (1961). "Molecular interpretation of the elasticity of resilin, a rubber-like protein". J. Mol. Biol. 3 (5): 648–667. doi:10.1016/s0022-2836(61)80028-4.
- Denny, M. W.; Gosline, J.M (1986). "The Structure and Properties of Spider Silk". Endeavor. 10: 37–43. doi:10.1016/0160-9327(86)90049-9.
- Alberts, Bruce (2002). "Molecular Biology of the Cell, 4th edition". Garland Science. 4: 1–43.
- Alberts, Bruce; Johnson, Alexander; Lewis, Julian; Raff, Martin; Roberts, Keith; Walter, Peter (2002-01-01). "The Plant Cell Wall". Cite journal requires
|journal=(help) - Hu, Shihao (2012). "Rational Design and Nanofabrication of Gecko-Inspired Fibrillar Adhesives". Nano Micro Small. 8 (16): 2464–2468. doi:10.1002/smll.201200413. PMID 22641471.