Body odour and sexual attraction
Odour is sensory stimulation of the olfactory membrane of the nose by a group of molecules.[1] Certain body odours are connected to human sexual attraction.[2][3] Humans can make use of body odour subconsciously to identify whether a potential mate will pass on favourable traits to their offspring. Body odour may provide significant cues about the genetic quality, health and reproductive success of a potential mate.[4] Body odour affects sexual attraction in a number of ways including through human biology, the menstrual cycle and fluctuating asymmetry. The olfactory membrane plays a role in smelling and subconsciously assessing another human's pheromones.[5] It also affects the sexual attraction of insects and mammals. The major histocompatibility complex genes are important for the immune system, and appear to play a role in sexual attraction via body odour. Studies have shown that body odor is strongly connected with heterosexual females. The women in the study ranked body odor as more important for attraction than “looks”. Humans may not simply depend on visual and verbal senses to be attracted to a possible partner/mate.[6]
Olfactory membrane
The olfactory membrane is a thick yellow/brown structure, about one inch square, located in the upper nasal cavity of the human nose. Made up of olfactory receptors and glands, the membrane is used as a tool to smell others' body odor and pheromones.[7] Chemicals that produce odour pass through the olfactory membrane to the olfactory bulbs, which contain biological receptors that detect the chemicals, and respond with an electrical signal transmitted to the brain by the olfactory nerves.[8] The olfactory membrane plays a large role in why humans are attracted to persons biologically rather than physically; this relates directly to the sense of smell and not physical appearance. Olfactory communication is common in all animals and in recent studies have shown that humans have this communication trait as well. This kind of communication happens subconsciously, and often influences a person's attraction to another.[5]
Pheromones
Pheromones are chemical messengers produced and emitted by the body that contribute significantly to interpersonal attraction.[5] The two types of pheromones include signal and primer, each playing a distinct role in human behavior. Signal pheromones act as attractants and repellents; they are classified as short term behavioral pheromones. Primer pheromones produce long term changes in human behavior and hormone production. The vomeronasal organ is used to detect the pheromones of others. Pheromones emitted from sweat glands play a role in sexual attraction, sexual repulsion, mother-infant bonding, and menstrual cycles.[5]
Human biology
There is a relationship between various aspects of human biology and genetics with sexual attraction. This includes the role of the major histocompatibility complex (MHC), the human leukocyte antigen (HLA) and their different heterozygotic variations. Such genetic factors may play a role in sexual selection.[9] Signalling odours in reproduction are called attractants; their function is to bring about successful mating.[1]
The MHC (major histocompatibility complex) is a group of genes essential for the immune system, playing an important role in immunological recognition.[10] These olfactory cues are involved in mate choice and preferences. HLA refers to the human form of MHC,[10] and is a gene complex which encodes the MHC.
MHC
There is a correlation between mate choice, odour preference and genetic similarity at the MHC.[10] Unique body smells are heavily influenced by MHC; these olfactory cues are probably involved in mate choice and preferences.[11]
MHC is expressed codominantly; a more diverse set of MHC genes leads to a stronger immune system.[12] Women prefer male partners with differing MHC genes from themselves. An evolutionary explanation is that females are attracted to males with MHC alleles different from their own, to provide their offspring with a stronger immune system.[13]
Females not using hormonal contraceptives were more attracted to the scent of males with dissimilar MHCs. Females currently using hormonal contraceptives preferred the scent of men with MHCs similar to their own.[14] The scent of an individual with low fluctuating asymmetry is universally more attractive.[15]
The increased attraction between people of dissimilar MHCs may help to prevent incest and subsequent birth defects.[16] This inbreeding avoidance hypothesis proposes that biases towards heterozygotic alleles prevent harmful genetic consequences which can arise from mating with genetic family members. Olfactory cues can be used to recognise kin.[17]
Couples who have many failed attempts at conception share a considerably larger amount of genes than those who are able to conceive without problems.[18] If, for these genetically similar couples, there is a successful conception, the babies are often born early or underweight.[19]
A third hypothesis explaining the function of dissimilar MHC mate preferences, the parasite hypothesis, suggests that MHC heterozygotes may be resistant to rapidly evolving parasites.[13]
HLA
The human leukocyte antigen system is a gene complex which encodes MHC in humans. Over thirty olfactory receptor genes have been located at the HLA class I region,[20] which presents peptides from inside the cell to be destroyed by the immune system. HLA- linked olfactory receptor genes can therefore provide a possible mechanism for detecting HLA- specific odours.[21]
Men and women are attracted to the pheromones they produce and HLA is related to the perceptions of other peoples' odours.[22] Males produce androstenol and androstenone. Androstenol is produced by fresh male sweat and is most attractive to women, while androstenone is produced once the sweat is exposed to oxygen and is highly unpleasant to women.
Studies have shown that women who are at the most fertile stage of their menstrual cycle prefer the smell of men that have higher testosterone levels.[23] Heterozygosity of HLA can also be detected through scent: in this case, heterozygosity confers greater ability to recognize a wider variety of antigens. Females, especially when not using hormonal contraceptives, are more attracted to the scent of males heterozygous for HLA.

However, the same attraction and mate preferences are not held by males for heterozygous females. Males are, however, more attracted to the scent of females with rare HLA alleles.[24] Men's arousal can be influenced by a variety of odours. For example; pumpkin pie, liquorice, doughnuts, and lavender can increase penile blood flow consequently causing arousal.[25]
Human pheromones and facial attraction
Two types of male pheromones: 5α-androst-16-en-3-one (MP1, androstenone) and 4,16-androstadien-3-one (MP2, androstadienone) and one type of female pheromone: 1,3,5(10),16-estratetrael-3-ol (FP, estratetraenol) are used as signals of mating quality. Studies[26] showed that MP2 has the strongest response produced by female vomeronasal organ (VNO), which is the first stage of the olfactory system. Sex-difference processing in the hypothalamus has been found between female and male pheromone, including in the VNO, where opposite-sex pheromones have different surface potential.[26][27][28] Studies have illustrated a relationship between human facial attraction and pheromones.[29] Individuals rated those of the opposite sex as more attractive if they preferred the individuals pheromones odour. It was also demonstrated that partners in a long-term relationship would have higher concordance between two types of signals of mating quality, vision and olfaction. In addition, no difference was found between two types of male pheromones in women's preference aside from the location of pheromones production.
Body odour and menstrual cycle changes
Women's fertility levels shift dramatically throughout the menstrual cycle, so the period surrounding ovulation is extremely important because it represents the peak period of reproductive fertility.[30] As conception is most likely to occur during a woman's brief fertile period, evolutionary theories suggest that men possess adaptations designed to maximize their reproductive success during this period.[30] Women's fertility shapes male mating behaviour, many studies have shown that being exposed to the scent of women's fertility led men to display greater implicit accessibility to mating-related concepts, males also judged the odours of women during the follicular phase as more pleasant and 'sexy' than odours during the luteal (non-ovulatory) phase.[3][30] Olfaction, therefore, transmits information relevant to human mate selection, through which men are capable of detecting shifts in women's fertility.[3][30]
There is now also considerable evidence from psychological studies that women's preferences for various male traits change throughout the menstrual cycle.[31] Hormonal fluctuation across the menstrual cycle explains temporal variation in women's judgment of the attractiveness of members of the opposite sex. This is due to the psychological processes that shape the formation and maintenance of human romantic relationships are influenced by variation in hormonal levels.[32]
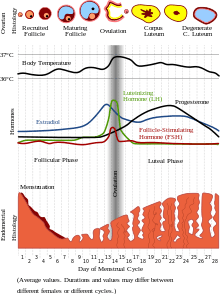
Due to their high hormone content, oral contraceptives have the potential to alter women's partner preference for a range of male traits, which could have important consequences on sexual relationships, as it alters women's attraction to their partner and, potentially, to other men.[31] If a woman's use of oral contraceptives is congruent, meaning she was using oral contraceptives when she met her partner, her current preference will more closely match the preference that shaped her partner choice in the first place, and the desire is higher than that of a woman whose use of oral contraceptives is incongruent.[31] The resulting factor is that women's partner preferences are influenced by oral contraceptives use, meaning that attraction towards an existing partner changes over time if a woman initiates or discontinues oral contraceptive use.[31]
Studies have shown that men are more attracted to women when they are more fertile and/or on their menstrual cycle. Women gave sample of when they were fertile and less fertile. The men in the study would smell and choose which sample between the two they were more attracted to. Women give off a more favorable smell the more fertile they are; in other words, men notice this and choose the more fertile sample rather than the less fertile sample.[33]
Body odour as a cue for ovulation
In contrast to previous literature, it has also been found that odours perceived from a female in follicular phase were actually more pleasant and sexier than odours perceived from that female in the luteal phase. Moreover, they discovered that the persistence of the menstrual cycle-dependent olfactory identification was extensive. However, the possibility of odours in the living environment overwhelming the menstrual cycle-dependent odours was not ruled out.[3] Repeat exposure to the specific odours increases the threshold level of odour detection and therefore reduce the stimulation in humans.[34] Additionally, it has been proposed through research that male in a relationship would be more sensitive to their partner's onset of ovulation-linked odour, and resultantly become desensitised after repeated exposure. On the other hand, non-paired males were continuously sensitive to the odour.[3] This demonstrates the persistence of the odour as an evolutionary or adaptation process, which could ensure the offspring reproduction success of female by keeping constant exposure to males during their menstrual cycle. During this period, the most fertile females tend to have more extra-pair copulations.[3][35] Males testosterone levels are found to be associated with a females’ reproduction fertility, such that males exposed to scents of females during the peak period of their fertility, had significantly higher testosterone levels than their baseline levels.[36] Evolution suggests this is an adaptation due to the physical changes inside the male body, although more research must be conducted to ensure these high levels of testosterone lead to reproductive behaviours.
Body odour and fluctuating asymmetry
Fluctuating Asymmetry (FA) is a type of biological asymmetry, referring to the extent to which small random deviations occur from expected perfect symmetry in different populations of organisms.[37] In humans, for example, FA can be demonstrated through the unequal sizes of bilateral features such as the eyes, ears and breasts. FA acts as an index for measuring developmental instability as it provides a clear indicator of the possible environmental and genetic stressors affecting development.[38] It is thought that having a preference for a symmetrical face offers some adaptive value as such symmetry may signal an individual's ability to cope with environmental challenges.[39] FA shares an inverse relationship with certain desired traits; a low FA is correlated with higher stress tolerance, larger body size in males, smaller body size in females, and higher facial attractiveness.[40] FA is detectable through the olfactory senses and it has a measurable effect on sexual attraction.[24] Significant cues may be found through body odour relating to a potential mate's health, reproductive status and genetic quality and FA is one such cue as it is considered to be a marker of genetic and developmental stability.[41]
During their fertile phase, females have repeatedly been shown as being more attracted to the body odours of more symmetrical males[42] and of males whose faces they rate as attractive.[43] The scent of symmetrical males provides an honest indicator of the male's phenotypic and genetic quality.[42] This may explain why women who are highly fertile find the scent of low FA as attractive and yet this scent is not necessarily as attractive to other women.[24] For example, it has been found that normally cycling women near their peak fertility tended to prefer the odour of shirts worn by symmetrical men and yet women at low fertility in their menstrual cycle or those using the contraceptive pill showed no preference for the odour of shirts of symmetrical men compared to those of asymmetrical men.[35] These findings support the good genes hypothesis such that when women are in the fertile phase of their menstrual cycle, they should prefer markers of genetic benefits or 'good genes'.[23] In other words, fertile women who prefer the scent of men with low FA are demonstrating a preference for the genetic benefits associated with those symmetric men.
It is widely accepted that men prefer the scent of women in her most fertile period. For example, in one study, men rated the body odours of T-shirts worn by women during their most fertile phase as more sexy and pleasant than T-shirts worn during their least fertile phase.[3] Despite this, much of the research in the area concludes that the effect of the scent of symmetry appears to be sex specific such that men do not find the scent of symmetrical women more attractive than the scent of asymmetrical women.[24][42] It therefore appears that attraction to symmetric body odour of the opposite sex appears to apply exclusively to women, and specifically fertile women, as non-fertile women and men do not display this preference.
Body odour in selection
Sex differences
.JPG)
Whilst a vast number of studies have been conducted to investigate body odour and mating, research is now shifting in particular towards the effect of male scent to female sexual attraction. This is largely due to the effects of the menstrual cycle and hormone contraceptives, which directly affect women's partner preference. Past research has highlighted the importance of a male's scent to females, such that smell was rated significantly more important for women than men. Furthermore, smell and body odour were rated as the most important physical factor for females, compared to looks for men.[44] Further studies have aimed to understand these sex differences. Using questionnaires and self-report data, a greater reliance was found for females on olfactory cues than males. This reliance was valued for females in both sexual and non-sexual contexts.[45] The research strongly supports the hypothesis that whilst men use more physical and visual cues, women rely more heavily on olfactory cues such as body odour that men tend to overlook.
Body odour enhancement
It has been established that women tend to rely more heavily on olfactory cues than males, rating those with pleasant body odour as more attractive than those with less pleasant body odour. Moreover, body odour and sexual attraction can be enhanced using artificial fragrances and dietary habits. Whilst males are influenced by body odour during selection, past research has demonstrated a significantly greater reliance on such cues by females.
A dependence on such olfactory cues has led to the enhancement of male body odour to influence female attraction. Past research has shown that videos of men using fragranced antiperspirants were rated as more attractive by women than those in a placebo control condition. In addition to this, females gave higher ratings of masculinity than males, particularly in the fragranced condition.[45] This provides evidence that females are more attentive to olfactory cues during partner selection. Aside from artificial fragrances, researchers have also begun to focus on more natural odour enhancements such as diet quality. A recent study aimed to investigate whether a dietary fruit and vegetable intake would influence female mate preferences. Its results showed that subjects rated as most pleasant smelling were those with a higher fruit and vegetable intake, suggesting male body odour can be enhanced by diet to appeal to females.[46]
Both sexes commonly use artificial scents to enhance their perceived sexual appeal. Many of these (musk compounds) share a similar chemical profile with naturally produced body chemicals.[47] Research has found a significant correlation between an individual's MHC genotype and the ratings they assigned to certain perfumes labelled to use "for self".[47] This therefore supports the hypothesis that perfumes can personally enhance body odours that indicate an individual's immunogenetics. However, several infertility issues may arise when people use perfumes or scented body washes that erase their natural scent, hindering women in particular from being able to detect if their partner is genetically comparable.
In animals
Insects
Insects use extremely sensitive receptors to detect pheromone signals. Each pheromone signal can elicit a distinct response based on the gender and social status of the recipient.[48] In insects, sex pheromones can be detected in very minute concentrations in the environment. Insect sex pheromones, usually released by the female to lure a male, are vital in the process by which insects locate each other for mating.[48] The main purpose of releasing these sex pheromones is to attract a partner from a distance, however the sex pheromones also serve to evoke a courtship response and sexually excite the male prior to copulation.[49] Male insects can also release sex pheromones, but this is only for the purpose of sexually exciting the female, making her more receptive to the male's advances. Generally, the majority of insects are sensitive and selective to the sex pheromone of their own species.
Insects make use of two classes of pheromone signals; the pheromones that induce immediate or releaser effects (for example, aggression or mating behaviours) and those that elicit long-lasting or ‘primer’ effects, such as physiological and hormonal changes.[48]

There is a significant amount of research supporting body odour and sexual attraction in insects. Observations and laboratory experiments of Culiseta inornata, identified a chemical substance involved in mating behavior, when exposed to this scent the male mosquitos were found to attempt sex with dead females and when exposed to the scent of virgin females, the males showed increased sexual activity through excited flight, searching and attempts to copulate with other males.[50] Further evidence comes from research on the commercial silkworm moth, Bombyx mori, a chemical produced in the abdominal sac of the female adult moth is released shortly after its emergence from the cocoon, male moths were found to be immediately attracted to this scent demonstrated by a characteristic wing flutter and attempts to copulate.[51] The sex pheromones of the silkworm moths can elicit responses in the male antenna at concentrations of only a few hundred molecules per square centimeter.[52]
Vertebrates
For vertebrates, aquatic environments are an ideal medium for dispersing chemical signals over large areas. Aquatic vertebrates use chemical signals for a wide range of purposes, from attracting mates to distant nesting sites during spawning, to signalling reproductive readiness and regulating predator/prey interactions.[48] Research on goldfish has identified that the fish release hormones in various combinations, depending on the reproductive status of the releaser, and these different combinations can elicit varying degrees of male courtship in the recipient.[53]
In terrestrial environments, chemosignals can be either volatile or non-volatile.[48] Accordingly, terrestrial vertebrates have two functionally and anatomically distinct olfactory systems: the main olfactory system, which is receptive to volatile cues, and the vomeronasal system, which is thought to process mostly non-volatile pheromones.[54]
Mammals
When it comes to sex, mammals use chemical signals (pheromones) to convey information to one another. Mammal's pheromones are air-borne chemical substances released in the urine or feces of animals or secreted from sweat glands that are perceived by the olfactory system and that elicit both behavioral and endocrine responses in conspecifics.[55] Mammals use sex pheromones to arouse, attract, and elicit specific behavioral responses from the opposite sex.[56] In mammals, chemical signals and the scent glands that secrete them have many features in common, for example, expression in only one sex, development only in adults, often only secreted in breeding season and used exclusively in mating.[57] For an odour to be exclusively result in sexual behavior, it must not only be perceived and preferred, but when absent there should be a decrease or complete elimination of sexual activity. This exclusivity has only been shown in golden hamsters[58] and the rhesus monkey.[59]
Mammalian pheromones can elicit both long-lasting effects that alter the hormone levels of the recipient animal, and short-term effects on its behavior.[60] For example, detection of male pheromones by female mice has been found to encourage onset of puberty, however the detection of female pheromones have been found to delay the onset of puberty.[61]
Odour can influence mammalian mating both directly and indirectly. Odour may act as a direct benefit to females, for example by avoiding contagious diseases by using odour cues to choose a healthy mate.[57] Odour can also act as an indicator mechanism, a form of indirect benefit, for example when a male displays a particular trait such as strength of odour which is in proportion to their heritable viability, females choosing males with strong odours will gain genes for high viability to pass to their offspring.[62]
There is vast evidence for the use of pheromones in mating behaviors. For example, when boars become sexually aroused, they salivate profusely dispersing pheromones into the air. These pheromones attract receptive sows, causing it to adopt a specific mating posture, known as standing, which allows the male boar to mount it and therefore copulate.[63]
Species specificity
Regardless of the species, sex pheromones are often structurally similar and for that reason different species need to be able to respond to the correct pheromone. It is the variation in the ratios of each compound within a pheromone that yields species specificity.[64] The use of mixtures of compounds as pheromones is well documented in insects, research into male orchid bees demonstrates that specific odours mediate exclusive attraction within a species.[65]
See also
References
- Michael, Stoddart, D. Michael (David) (1976-01-01). Mammalian odours and pheromones. Edward Arnold.
- Spehr, Marc; Kelliher, Kevin R.; Li, Xiao-Hong; Boehm, Thomas; Leinders-Zufall, Trese; Zufall, Frank (2006-02-15). "Essential Role of the Main Olfactory System in Social Recognition of Major Histocompatibility Complex Peptide Ligands". Journal of Neuroscience. 26 (7): 1961–1970. doi:10.1523/jneurosci.4939-05.2006. PMC 6674934. PMID 16481428.
- Singh, D.; Bronstad, P.M. (22 April 2001). "Female body odour is a potential cue to ovulation". Proceedings of the Royal Society B. 268 (1469): 797–801. doi:10.1098/rspb.2001.1589. PMC 1088671. PMID 11345323.
- Havlicek, Jan; Roberts, S. Craig; Flegr, Jaroslav (2005). "Women's preference for dominant male odour: effects of menstrual cycle and relationship status". Biology Letters. 1 (3): 256–259. doi:10.1098/rsbl.2005.0332. PMC 1617143. PMID 17148181.
- Grammer, Karl; Fink, Bernhard; Neave, Nick (February 2005). "Human pheromones and sexual attraction". European Journal of Obstetrics & Gynecology and Reproductive Biology. 118 (2): 135–142. doi:10.1016/j.ejogrb.2004.08.010. PMID 15653193.
- Herz, Rachel S.; Inzlicht, Michael (September 2002). "Sex differences in response to physical and social factors involved in human mate selection". Evolution and Human Behavior. 23 (5): 359–364. doi:10.1016/S1090-5138(02)00095-8.
- Nak-Eon Choi; Jung H. Han (23 February 2015). How Flavor Works: The Science of Taste and Aroma. John Wiley & Sons. p. 155. ISBN 978-1-118-86547-7.
- Brian R. Eggins (26 April 2002). Chemical Sensors and Biosensors. John Wiley & Sons. p. 3. ISBN 978-0-471-89914-3.
- Raphaëlle, C; Cao, C; Donnelly, P (2008). "Is Mate Choice in Humans MHC-Dependent?". PLoS Genet. 4 (9): e1000184. doi:10.1371/journal.pgen.1000184. PMC 2519788. PMID 18787687.
- Sandro Carvalho Santos, Pablo; Augusto Schinemann, Juliano; Gabardo, Juarez; da Graça Bicalho, Maria (2005-04-01). "New evidence that the MHC influences odor perception in humans: a study with 58 Southern Brazilian students". Hormones and Behavior. 47 (4): 384–388. doi:10.1016/j.yhbeh.2004.11.005. PMID 15777804.
- Breedlove, Marc; Watson, Neil (2013). Biological Psychology (Seventh Edition). Sunderland, Massachusetts: Sinauer Associates. p. 372. ISBN 978-0-87893-927-5.
- Janeway, Charles A; Travers, Paul; Walport, Mark; Shlomchik, Mark J (2001). Immunobiology (5 ed.). New York: Garland Science. pp. The major histocompatibility complex and its functions. ISBN 978-0-8153-3642-6.
- Penn, Dustin; Potts, Wayne (1999). "The Evolution of Mating Preferences and Major Histocompatibility Complex Genes". The American Naturalist. 154 (2): 145–164. CiteSeerX 10.1.1.616.9744. doi:10.1086/303166. JSTOR 101086/303166. PMID 29578757.
- Wedekind, Claus; Seebeck, Thomas; Bettens Florence; Paepke, Alexander J. (22 June 1995). "MHC-Dependent Mate Preferences in Humans". Proceedings of the Royal Society B. 260 (1359): 245–249. Bibcode:1995RSPSB.260..245W. doi:10.1098/rspb.1995.0087. PMID 7630893.
- Manning, J; Koukourakis, K; Brodie, D (1997). "Fluctuating asymmetry, metabolic rate and sexual selection in human males". Evolution and Human Biology. 18: 15–21. doi:10.1016/s1090-5138(96)00072-4.
- Potts, W; Manning, J; Wakeland, E; Hughes, A (1994). "The Role of Infectious Disease, Inbreeding and Mating Preferences in Maintaining MHC Genetic Diversity: An Experimental Test". Philosophical Transactions of the Royal Society of London B: Biological Sciences. 346 (1317): 369–378. doi:10.1098/rstb.1994.0154. PMID 7708831.
- Brown, Jerram L.; Eklund, Amy (1994-01-01). "Kin Recognition and the Major Histocompatibility Complex: An Integrative Review". The American Naturalist. 143 (3): 435–461. doi:10.1086/285612. JSTOR 2462739.
- Ober, Carole; Weitkamp, Lowell R.; Cox, Nancy; Dytch, Harvey; Kostyu, Donna; Elias, Sherman (1997-09-01). "HLA and Mate Choice in Humans". The American Journal of Human Genetics. 61 (3): 497–504. doi:10.1086/515511. PMC 1715964. PMID 9326314.
- Laitinen, Tarja (1993). "A Set of MHC Haplotypes Found Among Finnish Couples Suffering From Recurrent Spontaneous Abortions". American Journal of Reproductive Immunology. 29 (3): 148–154. doi:10.1111/j.1600-0897.1993.tb00580.x.
- Ehlers, Anke; Beck, Stephan; Forbes, Simon A.; Trowsdale, John; Volz, Armin; Younger, Ruth; Ziegler, Andreas (2000-12-01). "MHC-Linked Olfactory Receptor Loci Exhibit Polymorphism and Contribute to Extended HLA/OR-Haplotypes". Genome Research. 10 (12): 1968–1978. doi:10.1101/gr.10.12.1968. PMC 313068. PMID 11116091.
- Eklund, Amy C; Belchak, Mary Margaret; Lapidos, Karen; Raha-Chowdhury, Ruma; Ober, Carole (2000-07-01). "Polymorphisms in the HLA-linked olfactory receptor genes in the Hutterites". Human Immunology. 61 (7): 711–717. doi:10.1016/S0198-8859(00)00132-4. PMID 10880742.
- Santos, PS; Schinemann, JA; Gabardo, J; da Graça Bicalho, M (2005). "New evidence that the MHC influences odor perception in humans: a study with 58 Southern Brazilian students". Hormones and Behavior. 47 (4): 384–388. doi:10.1016/j.yhbeh.2004.11.005. PMID 15777804.
- Gangestad, S.W; Garver-Apgar, C.E; Simpson, J.A; Cousins, A.J (2007). "Changes in women's mate preferences across the ovulatory cycle". Journal of Personality and Social Psychology. 92 (1): 151–163. CiteSeerX 10.1.1.495.1474. doi:10.1037/0022-3514.92.1.151. PMID 17201549.
- Thornhill, R. (1 September 2003). "Major histocompatibility complex genes, symmetry, and body scent attractiveness in men and women". Behavioral Ecology. 14 (5): 668–678. doi:10.1093/beheco/arg043.
- Hirsch, A; Gruss, J (1999). "Human Male Sexual Response to Olfactory Stimuli". American Academy of Neurological and Orthopaedic Surgeons. 19: 14–19.
- Jennings-White, C (1995). "Perfumery and the sixth sense". Perfumer and Flavorist. 20: 1.
- Savic, Ivanka; Berglund, Hans; Gulyas, Balazs; Roland, Per (August 2001). "Smelling of Odorous Sex Hormone-like Compounds Causes Sex-Differentiated Hypothalamic Activations in Humans". Neuron. 31 (4): 661–668. doi:10.1016/s0896-6273(01)00390-7. PMID 11545724.
- Monti-Bloch, L.; Grosser, B.I. (October 1991). "Effect of putative pheromones on the electrical activity of the human vomeronasal organ and olfactory epithelium". The Journal of Steroid Biochemistry and Molecular Biology. 39 (4): 573–582. doi:10.1016/0960-0760(91)90255-4. PMID 1892788.
- Cornwell, R. Elisabeth; Boothroyd, Lynda; Burt, D. Michael; Feinberg, David R.; Jones, Ben C.; Little, Anthony C.; Pitman, Robert; Whiten, Susie; Perrett, David I. (2004-03-22). "Concordant preferences for opposite–sex signals? Human pheromones and facial characteristics". Proceedings of the Royal Society of London B: Biological Sciences. 271 (1539): 635–640. doi:10.1098/rspb.2003.2649. PMC 1691642. PMID 15156922.
- Miller, Saul L.; Maner, Jon K. (2011). "Ovulation as a male mating prime: Subtle signs of women's fertility influence men's mating cognition and behavior". Journal of Personality and Social Psychology. 100 (2): 295–308. doi:10.1037/a0020930. PMID 20822287.
- Roberts, S. Craig; Cobey, Kelly D.; Klapilová, Kateřina; Havlíček, Jan (2013-06-04). "An Evolutionary Approach Offers a Fresh Perspective on the Relationship Between Oral Contraception and Sexual Desire". Archives of Sexual Behavior. 42 (8): 1369–1375. doi:10.1007/s10508-013-0126-9. hdl:1893/18462. PMID 23733157.
- Roberts, S. Craig; Little, Anthony C.; Burriss, Robert P.; Cobey, Kelly D.; Klapilová, Kateřina; Havlíček, Jan; Jones, Benedict C.; DeBruine, Lisa; Petrie, Marion (2014-07-01). "Partner Choice, Relationship Satisfaction, and Oral Contraception The Congruency Hypothesis" (PDF). Psychological Science. 25 (7): 1497–1503. doi:10.1177/0956797614532295. PMID 24818612.
- Gildersleeve, Kelly A.; Haselton, Martie G.; Larson, Christina M.; Pillsworth, Elizabeth G. (February 2012). "Body odor attractiveness as a cue of impending ovulation in women: Evidence from a study using hormone-confirmed ovulation". Hormones and Behavior. 61 (2): 157–166. doi:10.1016/j.yhbeh.2011.11.005. PMID 22137971.
- Dalton, P. (2000-08-01). "Psychophysical and Behavioral Characteristics of Olfactory Adaptation". Chemical Senses. 25 (4): 487–492. doi:10.1093/chemse/25.4.487. PMID 10944515.
- Gangestad, Steven W.; Thornhill, Randy (1998-05-22). "Menstrual cycle variation in women's preferences for the scent of symmetrical men". Proceedings of the Royal Society of London B: Biological Sciences. 265 (1399): 927–933. doi:10.1098/rspb.1998.0380. PMC 1689051. PMID 9633114.
- Miller, Saul L.; Maner, Jon K. (2010-02-01). "Scent of a Woman Men's Testosterone Responses to Olfactory Ovulation Cues". Psychological Science. 21 (2): 276–283. doi:10.1177/0956797609357733. PMID 20424057.
- Tomkins, Joseph L; Kotiaho, Janne S (2002). Fluctuating asymmetry. eLS. doi:10.1038/npg.els.0003741. ISBN 978-0470016176.
- Møller, A.P.; Pomiankowski, A (1993). "Fluctuating asymmetry and sexual selection". Personality and Individual Differences. 89 (1–3): 267–279. doi:10.1007/BF02424520.
- Fink, B.; Neave, N.; Manning, J.T.; Grammer, K (2006). "Facial symmetry and judgements of attractiveness, health and personality". Personality and Individual Differences. 41 (3): 491–499. doi:10.1016/j.paid.2006.01.017.
- Manning, J.T.; Koukourakis, K.; Brodie, D.A. (1997). "Fluctuating asymmetry, metabolic rate and sexual selection in human males". Evolution and Human Biology. 18: 15–21. doi:10.1016/S1090-5138(96)00072-4.
- Havlicek, Jan; Roberts, S. Crag; Flegr, Jaroslav (2005). "Women's preference for dominant male odour: effects of menstrual cycle and relationship status". Biology Letters. 1 (3): 256–259. doi:10.1098/rsbl.2005.0332. PMC 1617143. PMID 17148181.
- Thornhill, Randy; Gangestad, Steven W. (1999). "The scent of symmetry: A human sex pheromone that signals fitness?". Evolution and Human Behavior. 20 (3): 175–201. doi:10.1016/S1090-5138(99)00005-7.
- Rikowski, Anya; Grammer, Karl (1999). "Human body odour, symmetry and attractiveness". Proceedings of the Royal Society B. 266 (1422): 869–874. doi:10.1098/rspb.1999.0717. PMC 1689917. PMID 10380676.
- Herz, Rachel S.; Inzlicht, Michael (2002). "Sex differences in response to physical and social factors involved in human mate selection". Evolution and Human Behavior. 23 (5): 359–364. doi:10.1016/s1090-5138(02)00095-8.
- Allen, Caroline; Cobey, Kelly D.; Havlíček, Jan; Roberts, S. Craig (2016). "The impact of artificial fragrances on the assessment of mate quality cues in body odor". Evolution and Human Behavior. 37 (6): 481–489. doi:10.1016/j.evolhumbehav.2016.05.001. hdl:1893/23162.
- Zuniga, Andrea; Stevenson, Richard J.; Mahmut, Mehmut K.; Stephen, Ian D. (2017). "Diet quality and the attractiveness of male body odor". Evolution and Human Behavior. 38 (1): 136–143. doi:10.1016/j.evolhumbehav.2016.08.002.
- Milinski, Manfred; Wedekind, Claus (2001-03-01). "Evidence for MHC-correlated perfume preferences in humans". Behavioral Ecology. 12 (2): 140–149. doi:10.1093/beheco/12.2.140.
- Dulac, Catherine; Torello, A. Thomas (July 2003). "Sensory systems: Molecular detection of pheromone signals in mammals: from genes to behaviour". Nature Reviews Neuroscience. 4 (7): 551–562. doi:10.1038/nrn1140. PMID 12838330.
- Jacobson, Martin (1972). Insect Sex Pheromones. New York: Academic Press. p. 1.
- Kliewer, J. W.; Miura, T.; Husbands, R. C.; Hurst, C. H. (1 May 1966). "Sex Pheromones and Mating Behavior of Culiseta inornata (Diptera: Culicidae)". Annals of the Entomological Society of America. 59 (3): 530–533. doi:10.1093/aesa/59.3.530.
- Butenandt, A; Beckmann, R; Stamm, D (1961). "Über den Sexuallockstoff des Seidenspinners, II. Konstitution und Konfiguration des Bombykols". Hoppe-Seyler's Zeitschrift für Physiologische Chemie. 324: 84–87. doi:10.1515/bchm2.1961.324.1.84.
- Wilson, Edward O. (1963). "Pheromones". Scientific American. 208 (5): 100–114. Bibcode:1963SciAm.208e.100W. doi:10.1038/scientificamerican0563-100.
- Poling, Kirsten R.; Fraser, E. Jane; Sorensen, Peter W. (2001-06-01). "The three steroidal components of the goldfish preovulatory pheromone signal evoke different behaviors in males". Comparative Biochemistry and Physiology B. 4TH International Symposium on Fish Endocrinology. 129 (2–3): 645–651. doi:10.1016/S1096-4959(01)00361-X.
- Broman, Prof Dr Ivar (1920-01-01). "Das Organon vomero-nasale Jacobsoni — ein Wassergeruchsorgan!". Anatomische Hefte (in German). 58 (1): 137–191. doi:10.1007/BF02033831.
- Rekwot, P.I.; Ogwu, D.; Oyedipe, E.O.; Sekoni, V.O. (March 2001). "The role of pheromones and biostimulation in animal reproduction". Animal Reproduction Science. 65 (3–4): 157–170. doi:10.1016/s0378-4320(00)00223-2.
- The Neurobiology of Olfaction. Boca Raton, FL: CRC Press/Taylor & Francis. 2009.
- Wyatt, Tristram D. (2003). Pheromones and animal behaviour : communication by smell and taste (Repr. with corrections 2004. ed.). Cambridge: Cambridge University Press. ISBN 9780521485265.
- Murphy, M. R.; Schneider, G. E. (16 January 1970). "Olfactory Bulb Removal Eliminates Mating Behavior in the Male Golden Hamster". Science. 167 (3916): 302–304. Bibcode:1970Sci...167..302M. doi:10.1126/science.167.3916.302.
- Michael, R. P.; Keverne, E. B.; Bonsall, R. W. (28 May 1971). "Pheromones: Isolation of Male Sex Attractants from a Female Primate". Science. 172 (3986): 964–966. Bibcode:1971Sci...172..964M. doi:10.1126/science.172.3986.964. PMID 4995585.
- Halpern, M (March 1987). "The Organization and Function of the Vomeronasal System". Annual Review of Neuroscience. 10 (1): 325–362. doi:10.1146/annurev.ne.10.030187.001545. PMID 3032065.
- Halpern, M. (1987-01-01). "The Organization and Function of the Vomeronasal System". Annual Review of Neuroscience. 10 (1): 325–362. doi:10.1146/annurev.ne.10.030187.001545. PMID 3032065.
- Andersson, Malte (1994). Sexual selection. Princeton, NJ: Princeton Univ. Press. ISBN 9780691000572.
- Dorries, Kathleen M.; Adkins-Regan, Elizabeth; Halpern, Bruce P. (1997). "Sensitivity and Behavioral Responses to the Pheromone Androstenone Are Not Mediated by the Vomeronasal Organ in Domestic Pigs". Brain, Behavior and Evolution. 49 (1): 53–62. doi:10.1159/000112981. PMID 8980852.
- Dobele, Angela; Lindgreen, Adam; Beverland, Michael; Vanhamme, Joëlle; van Wijk, Robert (2007-07-01). "Why pass on viral messages? Because they connect emotionally" (PDF). Business Horizons. 50 (4): 291–304. doi:10.1016/j.bushor.2007.01.004.
- Zimmermann, Yvonne; Roubik, David W.; Eltz, Thomas (19 July 2006). "Species-specific attraction to pheromonal analogues in orchid bees". Behavioral Ecology and Sociobiology. 60 (6): 833–843. doi:10.1007/s00265-006-0227-8.