TetR
Tet Repressor proteins (otherwise known as TetR) are proteins playing an important role in conferring antibiotic resistance to large categories of bacterial species.
.png)
Tetracycline (tc) is a broad family of antibiotics to which bacteria have evolved resistance. Tc normally kills bacteria by binding to the bacterial ribosome and halting protein synthesis. The expression of tc resistance genes is regulated by the repressor TetR. TetR represses the expression of TetA, a membrane protein that pumps out substances toxic to the bacteria like tc, by binding the tetA operator.[1] In tc resistant bacteria, TetA will pump out tc before it can bind to the ribosome because the repressive action of TetR on TetA is halted by binding of tc to TetR.[1] Therefore, TetR may have an important role in helping scientists to better understand mechanisms of antibiotic resistance and how to treat antibiotic resistant bacteria. TetR is one of many proteins in the TetR protein family, which is so named because TetR is the most well characterized member.[2]
TetR is used in artificially engineered gene regulatory networks because of its capacity for fine regulation of promoters. In the absence of tc or analogs like aTc, basal expression of TetR-regulated promoters is low, but expression rises sharply in the presence of even a minute quantity of tc. The tetA gene is also present in the widely used E. coli cloning vector pBR322, where it is often referred to by the name of its tetracycline-resistance phenotype, TetR, not to be confused with TetR.[3]
Structure & Function
.png)
TetR functions as a homodimer.[1] Each monomer consists of ten alpha helices connected by loops and turns. The overall structure of TetR can be broken down into two DNA-binding domains (one per monomer) and a regulatory core, which is responsible for tetracycline recognition and dimerization. TetR dimerizes by making hydrophobic contacts within the regulatory core. There is a binding cavity for tetracycline in the outer helices of the regulatory domain. When tetracycline binds this cavity, it causes a conformational change that affects the DNA-binding domain so that TetR is no longer able to bind DNA. As a result, TetA and TetR are expressed. There is still some debate in the field whether tetracycline derivatives alone can cause this conformational change or whether tetracycline must be in complex with magnesium to bind TetR.[4] (TetR typically binds tetracycline-Mg2+ complexes inside bacteria, but TetR binding to tetracycline alone has been observed in vitro.)
.png)
The DNA-binding domains of TetR recognize a 15 base pair palindromic sequence of the TetA operator.[1][5] These domains mainly consist of a helix-turn-helix (HTH) motif that is common in TetR protein family members (see below). However, the N-terminal residues preceding this motif have also been shown to be important for DNA binding.[6] Although these residues do not directly contact the DNA, they pack against the HTH and this packing is essential for binding. The HTH motifs have mostly hydrophobic interactions with major grooves of the target DNA.[1] Binding of TetR to its target DNA sequence causes changes in both the DNA and TetR.[7] TetR causes widening of the major grooves as well as kinking of the DNA; one helix of the HTH motif of TetR adopts a 310 helical turn as the result of complex DNA interactions.
TetR Protein Family
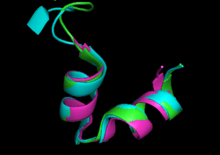
As of June 2005, this family of proteins had about 2,353 members that are transcriptional regulators.[1] (Transcriptional regulators control gene expression.) These proteins contain a helix-turn-helix (HTH) motif that is the DNA-binding domain. The second helix is considered to be most important for DNA sequence specificity and often recognizes nucleic acids within the major groove of the double helix.[7] In the majority of the family members, this motif is on the N-terminal end of the protein and is highly conserved.[1] The high conservation of the HTH motif is not observed for the other domains of the protein. The differences observed in these other regulatory domains are likely due to differences in the molecules that each family member senses.
TetR protein family members are mostly transcriptional repressors, meaning that they prevent the expression of certain genes at the DNA level. These proteins can act on genes with various functions including antibiotic resistance, biosynthesis and metabolism, bacterial pathogenesis, and response to cell stress.
See also
- Tetracycline controlled transcriptional activation
References
- Ramos JL, Martínez-Bueno M, Molina-Henares AJ, Terán W, Watanabe K, Zhang X, et al. (June 2005). "The TetR family of transcriptional repressors". Microbiology and Molecular Biology Reviews. 69 (2): 326–56. doi:10.1128/mmbr.69.2.326-356.2005. PMC 1197418. PMID 15944459.
- "InterPro". www.ebi.ac.uk. Retrieved 2020-08-06.
- Allard JD, Bertrand KP (September 1992). "Membrane topology of the pBR322 tetracycline resistance protein. TetA-PhoA gene fusions and implications for the mechanism of TetA membrane insertion". The Journal of Biological Chemistry. 267 (25): 17809–19. PMID 1517220.
- Werten S, Dalm D, Palm GJ, Grimm CC, Hinrichs W (December 2014). "Tetracycline repressor allostery does not depend on divalent metal recognition". Biochemistry. 53 (50): 7990–8. doi:10.1021/bi5012805. PMID 25432019.
- Orth P, Schnappinger D, Hillen W, Saenger W, Hinrichs W (March 2000). "Structural basis of gene regulation by the tetracycline inducible Tet repressor-operator system". Nature Structural Biology. 7 (3): 215–9. doi:10.1038/73324. PMID 10700280.
- Berens C, Altschmied L, Hillen W (January 1992). "The role of the N terminus in Tet repressor for tet operator binding determined by a mutational analysis". The Journal of Biological Chemistry. 267 (3): 1945–52. PMID 1309804.
- Huffman JL, Brennan RG (February 2002). "Prokaryotic transcription regulators: more than just the helix-turn-helix motif". Current Opinion in Structural Biology. 12 (1): 98–106. doi:10.1016/S0959-440X(02)00295-6. PMID 11839496.