Secotioid
Secotioid fungi are an intermediate growth form between mushroom-like hymenomycetes and closed bag-shaped gasteromycetes, where an evolutionary process of gasteromycetation has started but not run to completion. Secotioid fungi may or may not have opening caps, but in any case they often lack the vertical geotropic orientation of the hymenophore needed to allow the spores to be dispersed by wind, and the basidiospores are not forcibly discharged or otherwise prevented from being dispersed (e.g. gills completely inclosed and never exposed as in the secotioid form of Lentinus tigrinus)—note—some mycologists do not consider a species to be secotioid unless it has lost ballistospory.[1]

Explanation of secotioid development and gasteromycetation
Historically agarics and boletes (which bear their spores on a hymenium of gills or tubes respectively) were classified quite separately from the gasteroid fungi, such as puff-balls and truffles, of which the spores are formed in a large mass enclosed in an outer skin. However, in spite of this apparently very great difference in form, recent mycological research, both at microscopic[2] and molecular[3] level has shown that sometimes species of open mushrooms are much more closely related to particular species of gasteroid fungi than they are to each other. Fungi which do not open up to let their spores be dispersed in the air, but which show a clear morphological relation to agarics or boletes, constitute an intermediate form and are called secotioid.[2]
The word is derived from the name of the genus Secotium, which contains such species.[4]
On a microscopic scale, secotioid fungi do not expel their spores forcibly from the basidium; their spores are "statismospores". Like gasteroid fungi, secotioid species rely on animals such as rodents or insects to distribute their spores.
It can at times be disadvantageous for a mushroom to open up and free its spores in the usual way. If this development is aborted, a secotioid form arises, perhaps to be followed eventually by an evolutionary progression to a fully gasteroid form. This type of progression is called gasteromycetation and seems to have happened several times independently starting from various genera of "normal" mushrooms. This means that the secotioid and also the gasteroid fungi are polyphyletic. According to the paper by Thiers,[2] in certain climates and certain seasons, it may be an advantage to remain closed, because moisture can be conserved in that way.
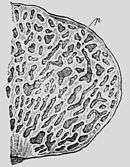
For example, the gasteroid genus Hymenogaster has been shown to be closely related to agaric genera such as Hebeloma, which were formerly placed in family Cortinariaceae or Strophariaceae. This is found by DNA analysis and also indicated on a microscopic scale by the resemblance of the spores and basidia. According to a current classification system, Hebeloma now belongs to family Hymenogastraceae, and is considered more narrowly related to the closed Hymenogaster fungi than, for instance, to the ordinary mushrooms in genus Cortinarius.[5][6]
A similar case is the well-known "Deceiver" mushroom Laccaria laccata which is now classified in the Hydnangiaceae, Hydnangium being a gastroid genus.
It has been found that a change in a single locus of a gene of the gilled mushroom Lentinus tigrinus causes it to have a closed fruiting body. This suggests that the emergence of a secotioid species may not require many mutations.[3]
There is a spectrum of secotioid species ranging from the open form to the closed form in the following respects:
- there may be an evident stipe, or there may be only a remnant consisting of a column of non-fertile tissue,
- if there is a stipe the edge of the cap may separate from it (partially opening), or may not,
- there may be recognizable gills (though oriented in all directions and very convoluted), or the fertile interior may be uniform like the gleba of gasteroid fungi, and
- the spore-bearing tissue may be above ground (epigeous), or underground (hypogeous), or partly buried.
The adjective "sequestrate" is sometimes used as a general term to mean "either secotioid or gasteroid".
Examples
Cortinarius is a very widespread genus of agarics, but also contains some secotioid species, such as C. leucocephalus, C. coneae and C. cartilagineus.
_Redhead_352483.jpg)
Pholiota nubigena is a secotioid species found early in the year at high altitude in the western United States. It was originally assigned to Secotium and later to a more specific secotioid genus Nivatogastrium, but in fact it is closely allied to Pholiota squarrosa[2] and it has now been moved to genus Pholiota itself, although the latter consists primarily of agarics.[7][8][9]{
Gastroboletus is a secotioid bolete genus where the fruiting bodies may or may not open, but in any case the tubes are not aligned vertically as in a true bolete.[2]

Agaricus deserticola is a secotioid species of Agaricus (the genus of common cultivated mushrooms) which at one time was placed in the genus Secotium. Similarly, Agaricus inapertus was formerly known as Endoptychum depressum until molecular analysis revealed it to be closely aligned with Agaricus.[10]
References
- Hibett DS; Tsuneda A; Shigeyuki M. (1994). "The Secotioid Form of Lentinus tegrinus:Genetics and Development of a Fungal Morphological Innovation". American Journal of Botany. 81 (4): 466–478. doi:10.2307/2445497. JSTOR 2445497.
- The following 1984 article provides a good introduction to sectioid fungi: Thiers, Harry D (January–February 1984). "The Secotioid Syndrome" (PDF). Mycologia. New York: The New York Botanical Garden, Bronx. NY 10458. 76 (1): 1–8. doi:10.2307/3792830. JSTOR 3792830.
- Hibbett, D. S; Pine, E. M; Langer, E; Langer, G; Donoghue, M. J (1997-10-28). "Evolution of gilled mushrooms and puffballs inferred from ribosomal DNA sequences". PNAS. Washington DC: National Academy of Sciences of the United States of America. 94 (22): 12002–12006. Bibcode:1997PNAS...9412002H. doi:10.1073/pnas.94.22.12002. PMC 23683. PMID 9342352.
- Conard, Henry S (March 1915). "The Structure and Development of Secotium agaricoides" (PDF). Mycologia. New York: The New York Botanical Garden, Bronx. NY 10458. 7 (2): 94–104. doi:10.2307/3753132. JSTOR 3753132.
- Binder, M; Bresinsky, A (2002). "Derivation of a polymorphic lineage of Gasteromycetes from boletoid ancestors". Mycologia. New York: The New York Botanical Garden, Bronx. NY 10458. 94 (1): 85–98. doi:10.2307/3761848. JSTOR 3761848. PMID 21156480.
- "Historical and current perspectives in the systematics of Australian cortinarioid sequestrate (truffle-like) fungi" (PDF). Australasian Mycologist. 21 (3): 81–116. 2002.
- See this Mykoweb page of Nivatogastrium nubigenum, which shows rudimentary lamellae.
- Matheny PB, Curtis JM, Hofstetter V, Aime MC, Moncalvo JM, Ge ZW, Yang ZL, Slot JC, Ammirati JF, Baroni TJ, Bougher NL, Hughes KW, Lodge DJ, Kerrigan RW, Seidl MT, Aanen DK, DeNitis M, Daniele GM, Desjardin DE, Kropp BR, Norvell LL, Parker A, Vellinga EC, Vilgalys R, Hibbett DS (2007). "Major clades of Agaricales: a multilocus phylogenetic overview". Mycologia. 98 (6): 982–95. doi:10.3852/mycologia.98.6.982. PMID 17486974.

- "the Pholiota nubigena page". Species Fungorum. Royal Botanic Gardens Kew. Retrieved 2020-05-24.
- Vellinga EC; de Kok RPJ; Bruns TD. (2003). "Phylogeny and taxonomy of Macrolepiota (Agaricaceae)". Mycologia. 95 (3): 442–56. doi:10.2307/3761886. JSTOR 3761886. PMID 21156633.