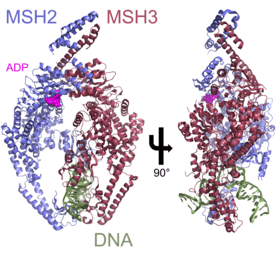
MSH3
DNA mismatch repair protein, MutS Homolog 3 (MSH3) is a human homologue of the bacterial mismatch repair protein MutS that participates in the mismatch repair (MMR) system. MSH3 typically forms the heterodimer MutSβ with MSH2 in order to correct long insertion/deletion loops and base-base mispairs in microsatellites during DNA synthesis. Deficient capacity for MMR is found in approximately 15% of colorectal cancers, and somatic mutations in the MSH3 gene can be found in nearly 50% of MMR-deficient colorectal cancers.[5]




Gene and Expression
In humans, the encoding gene for MSH3 is found on chromosome 5 at location 5q11-q12 upstream of the dihydrofolate reductase (DHFR) gene.[6][7] MSH3 is encoded by 222,341 base pairs and creates a protein consisting of 1137 amino acids.[8]
MSH3 is typically expressed at low levels in several transformed cell lines—including HeLa, K562, HL-60, and CEM—as well as a large range of normal tissues including spleen, thymus, prostate, testis, ovary, small intestine, colon, peripheral blood leukocytes, heart, brain, placenta, lung, liver, skeletal muscle kidney, and pancreas. Although expression levels of MSH3 vary slightly from tissue to tissue, its widespread low-level expression indicates that it is a “housekeeping” gene commonly expressed in all cells.[7]
Over-expression of MSH3 decreased capacity for MMR. When MSH3 is over expressed, drastic changes occur in the relative levels of formation of MutSβ at the expense of MutSα. MutSα is responsible for base-base mispairs and short insertion/deletion loops, while MutSβ repairs long insertion/deletion loops in DNA. A drastic shift in the relative levels of these protein complexes can lead to diminished capacity for MMR. In the case of MSH3 overexpression, MSH2 preferentially heterodimerizes with MSH3 leading to high levels of MutSβ and degradation of the partnerless MSH6 protein which normally complexes with MSH2 to form MutSα.[9]
Interactions
MSH3 has been shown to interact with MSH2, PCNA, and BRCA1. These interactions form protein complexes that are typically involved in tumor suppression and DNA repair activities.
The primary interaction of MSH3 involves forming the MutSβ complex with MSH2. MutSβ forms as a heterodimer of MSH2 and MSH3 with two primary interaction regions: an amino-terminal region and a carboxy-terminal region.[10] The N-terminal region of MSH3 (amino acids 126-250) contact the N-terminal region of MSH2 aa 378-625. The C-terminal regions connect at aa 1050-1128 of MSH3 and aa 875-934 of MSH2. The binding regions on MSH2 are identical when binding to either MSH3 or MSH6.[10] Adenine nucleotide binding regions in MSH3 and MSH2 are not contained in either of the interaction regions involved in dimerization, allowing MutSβ to bind to DNA and perform MMR.
{kind=link}
Proliferating cell nuclear antigen (PCNA) is a protein involved in post-replication MMR. It has been shown that PCNA binds to the MutSβ heterodimer via a binding motif in the N-terminal domain of MSH3. Bound PCNA then localizes the MutSβ complex to replication foci, indicating that PCNA assists in initiating repair by guiding MutSβ and other repair proteins to free termini in recently replicated DNA.[11]
Function
The primary function of MSH3 is to maintain the stability of the genome and enact tumor suppression by forming the heterodimer MutSβ to correct long insertion/deletion loops and base-base mispairs. In the case of long insertion/deletion loops, DNA is severely bent and downstream basepairs can become unpaired and exposed. MutSβ recognizes insertion/deletion loops of 1-15 nucleotides; binding to insertion/deletion loops is achieved by inserting the mismatch-binding domain of MSH3 and part of the mismatch-binding domain of MSH2 into the groove formed by the extreme bend in DNA formed by the insertion/deletion loop.[12]
{kind=link}
Role in Cancer
The most significant role of MSH3 in cancer is the suppression of tumors by repair of somatic mutations in DNA that occur as the result of base-base mispairs and insertion/deletion loops. Both loss of expression and over expression of MSH3 can lead to carcinogenic effects.
Over-expression of MSH3 can lead to drastic changes in the relative e levels of MutSα and MutSβ. Normally, MutSβ is expressed at relatively low levels throughout all cells while MutSα is present at high levels. While both proteins have redundant function in base-base repairs, MutSα typically effects base-base mispair repairs and also performs repairs on the more common short inertion/deletion loops. When MSH3 is heavily overexpressed, it acts as a sequester for MSH2 and the relative levels of MutSβ and MutSα shift dramatically as unpaired MSH6 proteins degrade and MutSα becomes depleted. MutSβ can compensate somewhat for loss of base-base mispair correction functions, but is not suited for repairing many short, 1-2 base pair insertion/deletion loops. This leads to a heightened rate of microsatellite instabilities and increased rates of somatic mutations.
This effect is directly related to human cancer in the form of drug resistance. One of the common resistance responses to methotrexate, a drug commonly used to treat childhood acute lymphocytic leukemia and a variety of other tumors, is amplification of the DHFR gene. DHFR amplification leads to overexpression of MSH3 and has been tied drug-resistant recurrence in cancer.[9]
In contrast, loss of MSH3 can lead to mismatch repair deficiency and genetic instability which have been identified as particularly common carcinogenic effects in human colorectal cancer. Mutations causing MSH3 knockdown can lead to diminished capacity for cells to repair long insertion/deletion loops causing microsatellite instabilities (MSI) in the genome and allowing an increase in the rates of somatic mutation. Elevated microsatellite alterations at selected tetranucleotide repeats (EMAST) are a type of MSI where loci containing AAAG or ATAG tetranucleotide repeats are particularly unstable. EMAST phenotypes are particularly common, with nearly 60% of sporadic colorectal cancers displaying high levels of EMAST linked to a high-rate of MSH3 deficient cells in tumors.[13]
References
- GRCh38: Ensembl release 89: ENSG00000113318 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000014850 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Gao, Jian-Xin; Park, Jae Myung; Huang, Shengbing; Tougeron, David; Sinicrope, Frank A. (2013). "MSH3 Mismatch Repair Protein Regulates Sensitivity to Cytotoxic Drugs and a Histone Deacetylase Inhibitor in Human Colon Carcinoma Cells". PLOS ONE. 8 (5): e65369. doi:10.1371/journal.pone.0065369. ISSN 1932-6203. PMC 3665625. PMID 23724141.
- MSH3 mutS homolog 3 [ Homo sapiens (human) ], retrieved 2014-05-10
- Watanabe A, Ikejima M, Suzuki N, Shimada T (1996). "Genomic organization and expression of the human MSH3 gene". Genomics. 31 (3): 311–8. doi:10.1006/geno.1996.0053. PMID 8838312.
- MutS Homolog 3 (Previous names: mutS (E. coli) homolog 3, mutS homolog 3 (E. coli)), retrieved 2014-05-10
- Marra G, Iaccarino I, Lettieri T, Roscilli G, Delmastro P, Jiricny J (1998). "Mismatch repair deficiency associated with overexpression of the MSH3 gene". Proc Natl Acad Sci U S A. 95 (15): 8568–73. doi:10.1073/pnas.95.15.8568. PMC 21116. PMID 9671718.
- Guerrette S, Wilson T, Gradia S, Fishel R (1998). "Interactions of human hMSH2 with hMSH3 and hMSH2 with hMSH6: examination of mutations found in hereditary nonpolyposis colorectal cancer". Mol Cell Biol. 18 (11): 6616–23. doi:10.1128/mcb.18.11.6616. PMC 109246. PMID 9774676.
- Kleczkowska HE, Marra G, Lettieri T, Jiricny J (2001). "hMSH3 and hMSH6 interact with PCNA and colocalize with it to replication foci". Genes Dev. 15 (6): 724–36. doi:10.1101/gad.191201. PMC 312660. PMID 11274057.
- Gupta S, Gellert M, Yang W (2012). "Mechanism of mismatch recognition revealed by human MutSβ bound to unpaired DNA loops". Nat Struct Mol Biol. 19 (1): 72–8. doi:10.1038/nsmb.2175. PMC 3252464. PMID 22179786.
- Haugen, A. C.; Goel, A.; Yamada, K.; Marra, G.; Nguyen, T.-P.; Nagasaka, T.; Kanazawa, S.; Koike, J.; Kikuchi, Y.; Zhong, X.; Arita, M.; Shibuya, K.; Oshimura, M.; Hemmi, H.; Boland, C. R.; Koi, M. (2008). "Genetic Instability Caused by Loss of MutS Homologue 3 in Human Colorectal Cancer". Cancer Research. 68 (20): 8465–8472. doi:10.1158/0008-5472.CAN-08-0002. ISSN 0008-5472. PMC 2678948. PMID 18922920.
Further reading
- Marti TM, Kunz C, Fleck O (2002). "DNA mismatch repair and mutation avoidance pathways". J. Cell. Physiol. 191 (1): 28–41. doi:10.1002/jcp.10077. PMID 11920679.
- Fujii H, Shimada T (1989). "Isolation and characterization of cDNA clones derived from the divergently transcribed gene in the region upstream from the human dihydrofolate reductase gene". J. Biol. Chem. 264 (17): 10057–64. PMID 2722860.
- Chen MJ, Shimada T, Moulton AD, et al. (1984). "The functional human dihydrofolate reductase gene". J. Biol. Chem. 259 (6): 3933–43. PMID 6323448.
- Shinya E, Shimada T (1994). "Identification of two initiator elements in the bidirectional promoter of the human dihydrofolate reductase and mismatch repair protein 1 genes". Nucleic Acids Res. 22 (11): 2143–9. doi:10.1093/nar/22.11.2143. PMC 308133. PMID 8029024.
- Risinger JI, Umar A, Boyd J, et al. (1996). "Mutation of MSH3 in endometrial cancer and evidence for its functional role in heteroduplex repair". Nat. Genet. 14 (1): 102–5. doi:10.1038/ng0996-102. PMID 8782829.
- Watanabe A, Ikejima M, Suzuki N, Shimada T (1997). "Genomic organization and expression of the human MSH3 gene". Genomics. 31 (3): 311–8. doi:10.1006/geno.1996.0053. PMID 8838312.
- Nakajima E, Orimo H, Ikejima M, Shimada T (1996). "Nine-bp repeat polymorphism in exon 1 of the hMSH3 gene". Jpn. J. Hum. Genet. 40 (4): 343–5. doi:10.1007/BF01900603. PMID 8851770.
- Acharya S, Wilson T, Gradia S, et al. (1997). "hMSH2 forms specific mispair-binding complexes with hMSH3 and hMSH6". Proc. Natl. Acad. Sci. U.S.A. 93 (24): 13629–34. doi:10.1073/pnas.93.24.13629. PMC 19374. PMID 8942985.
- Guerrette S, Wilson T, Gradia S, Fishel R (1998). "Interactions of human hMSH2 with hMSH3 and hMSH2 with hMSH6: examination of mutations found in hereditary nonpolyposis colorectal cancer". Mol. Cell. Biol. 18 (11): 6616–23. doi:10.1128/mcb.18.11.6616. PMC 109246. PMID 9774676.
- Ceccotti S, Ciotta C, Fronza G, et al. (2000). "Multiple mutations and frameshifts are the hallmark of defective hPMS2 in pZ189-transfected human tumor cells". Nucleic Acids Res. 28 (13): 2577–84. doi:10.1093/nar/28.13.2577. PMC 102707. PMID 10871409.
- Orimo H, Nakajima E, Yamamoto M, et al. (2000). "Association between single nucleotide polymorphisms in the hMSH3 gene and sporadic colon cancer with microsatellite instability". J. Hum. Genet. 45 (4): 228–30. doi:10.1007/s100380070031. PMID 10944853.
- Clark AB, Valle F, Drotschmann K, et al. (2001). "Functional interaction of proliferating cell nuclear antigen with MSH2-MSH6 and MSH2-MSH3 complexes". J. Biol. Chem. 275 (47): 36498–501. doi:10.1074/jbc.C000513200. PMID 11005803.
- Kleczkowska HE, Marra G, Lettieri T, Jiricny J (2001). "hMSH3 and hMSH6 interact with PCNA and colocalize with it to replication foci". Genes Dev. 15 (6): 724–36. doi:10.1101/gad.191201. PMC 312660. PMID 11274057.
- Schmutte C, Sadoff MM, Shim KS, et al. (2001). "The interaction of DNA mismatch repair proteins with human exonuclease I." J. Biol. Chem. 276 (35): 33011–8. doi:10.1074/jbc.M102670200. PMID 11427529.
- Wang Q, Zhang H, Guerrette S, et al. (2001). "Adenosine nucleotide modulates the physical interaction between hMSH2 and BRCA1". Oncogene. 20 (34): 4640–9. doi:10.1038/sj.onc.1204625. PMID 11498787.
- Mazurek A, Berardini M, Fishel R (2002). "Activation of human MutS homologs by 8-oxo-guanine DNA damage". J. Biol. Chem. 277 (10): 8260–6. doi:10.1074/jbc.M111269200. PMID 11756455.
- Plotz G, Raedle J, Brieger A, et al. (2002). "hMutSalpha forms an ATP-dependent complex with hMutLalpha and hMutLbeta on DNA". Nucleic Acids Res. 30 (3): 711–8. doi:10.1093/nar/30.3.711. PMC 100294. PMID 11809883.
- Arzimanoglou II, Hansen LL, Chong D, et al. (2002). "Frequent LOH at hMLH1, a highly variable SNP in hMSH3, and negligible coding instability in ovarian cancer". Anticancer Res. 22 (2A): 969–75. PMID 12014680.
- Ohta S, Shiomi Y, Sugimoto K, et al. (2002). "A proteomics approach to identify proliferating cell nuclear antigen (PCNA)-binding proteins in human cell lysates. Identification of the human CHL12/RFCs2-5 complex as a novel PCNA-binding protein". J. Biol. Chem. 277 (43): 40362–7. doi:10.1074/jbc.M206194200. PMID 12171929.