Hairpin ribozyme
The hairpin ribozyme is a small section of RNA that can act as a ribozyme. Like the hammerhead ribozyme it is found in RNA satellites of plant viruses. It was first identified in the minus strand of the tobacco ringspot virus (TRSV) satellite RNA where it catalyzes self-cleavage and joining (ligation) reactions to process the products of rolling circle virus replication into linear and circular satellite RNA molecules. The hairpin ribozyme is similar to the hammerhead ribozyme in that it does not require a metal ion for the reaction.
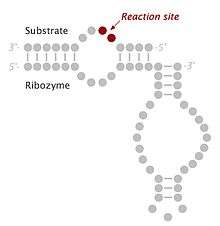
Biological function
The hairpin ribozyme is an RNA motif that catalyzes RNA processing reactions essential for replication of the satellite RNA molecules in which it is embedded. These reactions are self-processing, i.e. a molecule rearranging its own structure. Both cleavage and end joining reactions are mediated by the ribozyme motif, leading to a mixture of interconvertible linear and circular satellite RNA molecules. These reactions are important for processing the large multimeric RNA molecules that are generated by rolling circle replication. At the end of the replication cycle, these large intermediates of satellite RNA replication are processed down to unit length molecules (circular or linear) before they can be packaged by viruses and carried to other cells for further rounds of replication.[1]
Natural and artificial versions of the hairpin ribozyme
The hairpin ribozyme has been identified in only 3 naturally occurring sequences:
- satellite RNA of tobacco ringspot virus (sTRSV)[3][4]
- satellite RNA of chicory yellow mottle virus (sCYMV)[5]
- satellite RNA of arabis mosaic virus (sARMV)[6]
Smaller artificial versions of the hairpin ribozyme have been developed to enable a more detailed experimental analysis of the molecule.[7] This is a commonly used strategy for separating those parts of a self-processing RNA molecule that are essential for the RNA processing reactions from those parts which serve unrelated functions. Through this process, a 50 nucleotide minimal catalytic domain and a 14 nucleotide substrate were identified.[8] Using these artificially derived sequences, a trans-acting ribozyme was developed that can catalyze the cleavage of multiple substrate molecules. This strategy was important in that it allowed investigators to (i) apply biochemical methods for enzymatic analysis, (ii) conduct experiments to identify essential structural elements of the ribozyme-substrate complex, and (iii) develop engineered ribozymes that have been used for biomedical applications, including preventing the replication of pathogenic viruses, and the study of the function of individual genes.
Reaction chemistry
In common with several other ribozymes and protein ribonucleases, the cleavage reaction of the hairpin ribozyme generates RNA fragments with termini consisting of a 2',3'-cyclic phosphate and a 5'-hydroxyl group. The ligation reaction appears to be a simple reversal of cleavage, i.e. covalent joining of RNA fragments ending with a 2',3'-cyclic phosphate and a 5'-hydroxyl group to generate the ordinary 3'-5' phosphodiester linkage used in both RNA and DNA.
Studies of this reaction in multiple ribozymes have served to establish that the reaction chemistry (catalytic mechanism) is an endogenous property of the RNA molecule itself and is not mediated by metal ions, as is true for some protein enzymes and some other ribozymes.[9] Moreover, cleavage activity is still observed when Mg2+ is replaced by [Co(NH3)6]3+.[10] Co3+ binds NH3 so tightly in solution that NH3 does not dissociate to any appreciable extent, and therefore does not become protonated. This suggests there is no metal-catalyzed proton transfer or direct coordination to the RNA, but instead metals are only required for folding. Furthermore, in crystal structures of a ribozyme-inhibitor complex and a transition state mimic, it was shown that the three-dimensional architecture splays apart A-1 and G+1, positioning the 2'-OH of A-1 for an in-line nucleophilic attack on the scissile phosphate linkage. Additionally, G8, A38, and A9 have been suggested to play roles in the catalysis by deprotonating the 2'-OH of A-1, stabilizing the developing negative charge of the pentacoordinate phosphate oxygens, and protonating the 5'-O leaving group of G+1.[11][12]
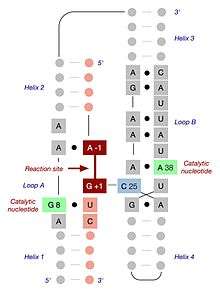
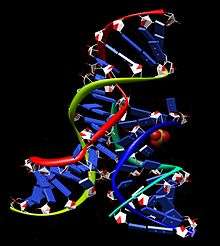
Structure
The minimal hairpin ribozyme-substrate complex folds into a secondary structure that includes two domains, each consisting of two short base paired helices separated by an internal loop. Domain A (helix 1 – loop A – helix 2) contains the substrate and the primary substrate-recognition region of the ribozyme. Domain B (helix 3 – loop B – helix 4) is larger and contains the primary catalytic determinants of the ribozyme. The two domains are covalently joined via a phosphodiester linkage that connects helix 2 to helix 3. These domains must interact with one another in order for catalysis to occur.[14]
When the minimal ribozyme-substrate complex is allowed to fold under conditions of low ionic strength, the two domains stack one atop the other, forming an inactive, extended structure that resembles a hairpin.[15] In order for catalysis to occur, the two domains lie parallel to one another in a fold that resembles a paperclip. In various publications, this RNA has been termed either the "paperclip" or "hairpin" ribozyme. Despite the fact that the former name has proven to be more accurate, the latter has become the commonly accepted nomenclature. In the laboratory, a functional interaction between the two domains is promoted by the addition of cations, whose positive charge suffices to overcome the electrostatic repulsion of the negatively charged RNA backbone. In nature, association of the two domains is assisted through a combination of metal ions (including Mg2+) and the presence of two additional helical domains that are not present in the minimal ribozyme-substrate complex but serve to promote proper three-dimensional folding. These additional domains stack upon helices 2 and 3, thereby promoting the association of the two functional domains through what is termed a four-way helical junction.[16]
The structure and activity of the hairpin ribozyme has been explored using a wide range of complementary experimental methods, including nucleotide replacement, functional group substitution, combinatorial selection, fluorescence spectroscopy, covalent crosslinking, NMR analysis and x-ray crystallography. These studies have been facilitated by the ability of the functional complex to self-assemble from segments made by solid phase chemical RNA synthesis, permitting the incorporation of a wide variety of modified nucleotides that are not naturally found in RNA. Together, the results of these experiments present a highly congruent picture of the catalytic cycle, i.e. how the hairpin ribozyme binds its substrate, folds into a specific three-dimensional structure, catalyzes the reaction, and releases the product(s) of the reaction.[17]
Targeted RNA cleavage and antiviral activity
Hairpin ribozymes have been modified in such a way that they can be used to target cleavage of other RNA molecules. This is possible because much of the substrate specificity of the hairpin ribozyme results from simple Watson-Crick base pairing within helices 1 and 2.[18]
One area of particular interest has been the development of hairpin ribozymes for potential therapeutic use, for example by preventing the replication of pathogenic viruses. Antiviral hairpin ribozymes have been generated and expressed within mammalian cells, and cells expressing different engineered ribozymes have been shown to be resistant to infection by HIV-1,[19][20] hepatitis B,[21] and Sindbis virus.[22]
References
- Symons, RH (1997). "Plant pathogenic RNAs and RNA catalysis". Nucleic Acids Res. 25 (14): 2683–2689. doi:10.1093/nar/25.14.2683. PMC 146833. PMID 9207012.
- Alam, S.; Grum-Tokars, V.; Krucinska, J.; Kundracik, M.; Wedekind, J. (2005). "Conformational heterogeneity at position U37 of an all-RNA hairpin ribozyme with implications for metal binding and the catalytic structure of the S-turn". Biochemistry. 44 (44): 14396–14408. doi:10.1021/bi051550i. PMID 16262240.
- Feldstein, PA; Buzayan, JM; Bruening, G (Oct 15, 1989). "Two sequences participating in the autolytic processing of satellite tobacco ringspot virus complementary RNA". Gene. 82 (1): 53–61. doi:10.1016/0378-1119(89)90029-2. PMID 2583519.
- Hampel, A; Tritz, R (Jun 13, 1989). "RNA catalytic properties of the minimum (-)sTRSV sequence". Biochemistry. 28 (12): 4929–4933. doi:10.1021/bi00438a002. PMID 2765519.
- Rubino, L; Tousignant, ME; Steger, G; Kaper, JM (September 1990). "Nucleotide sequence and structural analysis of two satellite RNAs associated with chicory yellow mottle virus". The Journal of General Virology. 71 (9): 1897–1903. doi:10.1099/0022-1317-71-9-1897. PMID 1698918.
- Kaper, JM; Tousignant, ME; Steger, G (Jul 15, 1988). "Nucleotide sequence predicts circularity and self-cleavage of 300-ribonucleotide satellite of arabis mosaic virus". Biochemical and Biophysical Research Communications. 154 (1): 318–325. doi:10.1016/0006-291x(88)90687-0. PMID 3395334.
- Feldstein, PA; Bruening, G (1993). "Catalytically active geometry in the reversible circularization of 'mini-monomer' RNAs derived from the complementary strand of tobacco ringspot virus satellite RNA". Nucleic Acids Research. 21 (8): 1991–1998. doi:10.1093/nar/21.8.1991. PMC 309442. PMID 7684131.
- Hampel, A; Tritz, R (1989). "RNA catalytic properties of the minimum (-)sTRSV sequence". Biochemistry. 28 (12): 4929–4933. doi:10.1021/bi00438a002. PMID 2765519.
- Murray, J. B.; Seyhan, A. A.; Walter, N. G.; Burke, J. M.; Scott, W. G. (1998). "The hammerhead, hairpin and VS ribozymes are catalytically proficient in monovalent cations alone" (PDF). Chemistry & Biology. 5 (10): 587–595. doi:10.1016/S1074-5521(98)90116-8. PMID 9818150.
- Young, K. J.; Gill, F.; Grasby, J. A. (1997-10-01). "Metal Ions Play a Passive Role in the Hairpin Ribozyme Catalysed Reaction". Nucleic Acids Research. 25 (19): 3760–3766. doi:10.1093/nar/25.19.3760. ISSN 0305-1048. PMC 146958. PMID 9380495.
- Rupert, P. B.; Ferré-D'Amaré, A. R. (2001-04-12). "Crystal structure of a hairpin ribozyme-inhibitor complex with implications for catalysis". Nature. 410 (6830): 780–786. Bibcode:2001Natur.410..780R. doi:10.1038/35071009. ISSN 0028-0836. PMID 11298439.
- Rupert, Peter B.; Massey, Archna P.; Sigurdsson, Snorri Th; Ferré-D'Amaré, Adrian R. (2002-11-15). "Transition state stabilization by a catalytic RNA". Science. 298 (5597): 1421–1424. Bibcode:2002Sci...298.1421R. doi:10.1126/science.1076093. ISSN 1095-9203. PMID 12376595.
- Salter, J.; Krucinska, J.; Alam, S.; Grum-Tokars, V.; Wedekind, J. E. (2006). "Water in the active site of an all-RNA hairpin ribozyme and the effects of Gua8 base variants on the geometry of phosphoryl transfer". Biochemistry. 45 (3): 686–700. doi:10.1021/bi051887k. PMC 2546605. PMID 16411744.
- Butcher, S. E.; Heckman, J. E.; Burke, J. M. (1995). "Reconstitution of hairpin ribozyme activity following separation of functional domains". The Journal of Biological Chemistry. 270 (50): 29648–29651. doi:10.1074/jbc.270.50.29648. PMID 8530348.
- Esteban, J. A.; Walter, N. G.; Kotzorek, G.; Heckman, J. E.; Burke, J. M. (1998). "Structural basis for heterogeneous kinetics: Reengineering the hairpin ribozyme" (PDF). Proceedings of the National Academy of Sciences of the United States of America. 95 (11): 6091–6096. Bibcode:1998PNAS...95.6091E. doi:10.1073/pnas.95.11.6091. PMC 27590. PMID 9600922.
- Nahas, M.; Wilson, T. J.; Ha, T.; Lilley, D. M. J. (2005). "Folding and catalysis of the hairpin ribozyme". Biochemical Society Transactions. 33 (Pt 3): 461–465. doi:10.1042/BST0330461. PMID 15916541.
- Cochrane, J. C.; Strobel, S. A. (2008). "Catalytic Strategies of Self-Cleaving Ribozymes". Accounts of Chemical Research. 41 (8): 1027–1035. doi:10.1021/ar800050c. PMID 18652494.
- Chowrira, BM; Burke, JM (1991). "Binding and cleavage of nucleic acids by the "hairpin" ribozyme". Biochemistry. 30 (35): 8518–8522. doi:10.1021/bi00099a003. PMID 1909564.
- Yu, M; Ojwang, J; Yamada, O; Hampel, A; Rapapport, J; Looney, D; Wong-Staal, F (1993). "A hairpin ribozyme inhibits expression of diverse strains of human immunodeficiency virus type 1". Proceedings of the National Academy of Sciences of the United States of America. 90 (13): 6340–6344. Bibcode:1993PNAS...90.6340Y. doi:10.1073/pnas.90.13.6340. PMC 46924. PMID 8327516.
- Puerta-Fernández E, Barroso-del Jesus A, Romero-López C, Tapia N, Martínez MA, Berzal-Herranz A (Jun 10, 2005). "Inhibition of HIV-1 replication by RNA targeted against the LTR region". AIDS. 19 (9): 863–870. doi:10.1097/01.aids.0000171399.77500.e0. PMID 15905666.
- Zu Putlitz, J.; Yu, Q.; Burke, J. M.; Wands, J. R. (1999). "Combinatorial Screening and Intracellular Antiviral Activity of Hairpin Ribozymes Directed against Hepatitis B Virus". Journal of Virology. 73 (7): 5381–5387. doi:10.1128/JVI.73.7.5381-5387.1999. PMC 112594. PMID 10364285.
- Zhang, Z.; Burke, J. (2005). "Inhibition of viral replication by ribozyme: mutational analysis of the site and mechanism of antiviral activity". Journal of Virology. 79 (6): 3728–3736. doi:10.1128/JVI.79.6.3728-3736.2005. PMC 1075678. PMID 15731266.
Further reading
- Doherty, EA; Doudna JA (2001). "Ribozyme structures and mechanisms". Annu Rev Biophys Biomol Struct. 30: 457–475. doi:10.1146/annurev.biophys.30.1.457. PMID 11441810.
- Ferré-D'Amaré, AR; Rupert PB (2002). "The hairpin ribozyme: from crystal structure to function". Biochem Soc Trans. 30 (Pt 6): 1105–1109. doi:10.1042/BST0301105. PMID 12440983.