Epstein–Barr virus stable intronic-sequence RNAs
Epstein–Barr virus stable intronic-sequence RNAs (ebv-sisRNAs) are a class of non-coding RNAs generated by repeat introns in the Epstein–Barr virus. After EBERs 1 and 2, ebv-sisRNA-1 is the third most abundant EBV RNA generated during a highly oncogenic form of virus latency (latency III). Conservation of ebv-sisRNA sequence and secondary structure between EBV and other herpesviruses suggest shared functions in latent infection.
| Epstein–Barr virus stable intronic sequence RNA | |
|---|---|
| Identifiers | |
| Symbol | EBVSIS |
| Other data | |
| RNA type | sisRNA |
| Domain(s) | Herpesviridae; |
| PDB structures | PDBe |
Background
The Epstein–Barr virus (EBV) infects as many as 95% of adults[1] and is the infectious agent responsible for mononucleosis ("mono"). Infection with EBV results in lifelong. Latent infections are "dormant", meaning no active virions are produced, however the virus generates proteins and RNAs to modulate host-virus interactions that maintain latent infection. In ways yet to be fully determined, these interactions make EBV-infected B cells more prone to becoming cancerous (e.g. Hodgkin's lymphoma, Burkitt's lymphoma, and nasopharyngeal carcinoma). Non-coding RNAs (ncRNAs) have a role in this process. Structured ncRNAs are of particular interest as they serve a wide array of functions, which are the focus of intensive efforts to characterize and archive in such projects as Rfam.[2][3]
A recent study of ncRNAs in EBV using bioinformatics and RNA-Seq identified multiple regions within its genome likely to contain functional RNAs.[4] These regions included EBER-1 and -2, v-snoRNA1, and most of the known viral miRNAs. In addition to these known EBV ncRNAs, this analysis identified new RNAs, including two stable intronic sequence (sis)RNAs. Introns, typically are rapidly degraded in the cell, but can persist and accumulate to high abundance when they serve a functional role. Such sisRNAs have been found in Xenopus oocytes.[5] Stable introns are also found in other herpesviruses, for example, the HHV Latency Associated Transcript, which plays important roles in the maintenance of virus latency.[6]

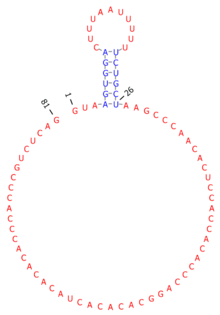
In EBV, sisRNAs are generated from a region known as the W repeats. This region is transcribed during a type of viral latency that is highly oncogenic (latency type III) and also in a rare type of latency (Wp-restricted latency) observed in ~15% of endemic Burkitt's lymphoma.[7] Splicing of these W repeat transcripts produces a short intron and a long intron (Fig. 1), both of which accumulate to high abundance in EBV-infected human B cells. Indeed, ebv-sisRNA-1 is the third most abundantly produced EBV RNA after EBER1 and EBER2, which are highly expressed in EBV-infected cells.,[8][9] The presence of these RNAs in a pathogenic form of latency suggests roles in EBV-associated cancers.[4]
ebv-sisRNA-1
The short W repeat intron, rather than being spliced and rapidly degraded, persists after splicing and is the third most abundant EBV-produced small ncRNA in latency III. Nucleotides 4 to 26 of ebv-sisRNA-1 form a short hairpin loop that presents a Uridine-rich sequence motif (a possible platform for protein interactions) into the loop. The remainder of the sequence is unlikely to form stable RNA structure. This unstructured stretch of sequence may be exposed to allow for interactions with nucleic acids or other proteins. The sisRNA sequence is ~100% conserved in EBV strains and homology extends to include other lymphocryptoviruses. The hairpin structure is also conserved and includes structure-preserving mutations in its stem.[5]
ebv-sisRNA-2
Ebv-sisRNA-2 is generated from the long W repeat intron. Evidence of stable and conserved RNA structure covers ~40% of this RNA and one region can fold into a remarkably long (586 nt) and thermodynamically stable hairpin loop (Fig. 2). In addition to EBV strains, where the hairpin is ~100% conserved in sequence, this structure is also found in other lymphocrypoviruses. Despite high divergence of sequence between these homologous RNAs, the long hairpin structure is well conserved. This suggests that this RNA plays an important functional role in ebv-sisRNA-2.[5] The size of ebv-sisRNA-2 (2,791 nt) more closely resembles that the HHV Latency Associated Transcript) and may perhaps play a similar role in the maintenance of virus latency.[6]
References
- "Epstein–Barr Virus and Infectious Mononucleosis". Archived from the original on 20 April 2012. Retrieved 11 November 2013.
- Heilman, J. M.; Kemmann, E.; Bonert, M.; Chatterjee, A.; Ragar, B.; Beards, G. M.; Iberri, D. J.; Harvey, M.; Thomas, B.; Stomp, W.; Martone, M. F.; Lodge, D. J.; Vondracek, A.; De Wolff, J. F.; Liber, C.; Grover, S. C.; Vickers, T. J.; Meskó, B.; Laurent, M. L. R. (2011). "Wikipedia: A Key Tool for Global Public Health Promotion". Journal of Medical Internet Research. 13 (1): e14. doi:10.2196/jmir.1589. PMC 3221335. PMID 21282098.
- Daub, J.; Gardner, P. P.; Tate, J.; Ramsköld, D.; Manske, M.; Scott, W. G.; Weinberg, Z.; Griffiths-Jones, S.; Bateman, A. (2008). "The RNA WikiProject: Community annotation of RNA families". RNA. 14 (12): 2462–2464. doi:10.1261/rna.1200508. PMC 2590952. PMID 18945806.
- Moss WN, Steitz JA (August 2013). "Genome-wide analyses of Epstein–Barr virus reveal conserved RNA structures and a novel stable intronic sequence RNA". BMC Genomics. 14: 543. doi:10.1186/1471-2164-14-543. PMC 3751371. PMID 23937650.
- Gardner EJ, Nizami ZF, Talbot CC, Gall JG (2012). "Stable intronic sequence RNA (sisRNA), a new class of noncoding RNA from the oocyte nucleus of Xenopus tropicalis". Genes & Development. 26 (22): 2550. doi:10.1101/gad.202184.112. PMC 3505824. PMID 23154985.
- Farrell MJ, Dobson AT, Feldman LT (1991-02-01). "Herpes simplex virus latency-associated transcript is a stable intron". Proceedings of the National Academy of Sciences. 88 (3): 790–794. doi:10.1073/pnas.88.3.790. PMC 50899. PMID 1846963. (Free full-text article: )
- Kelly, G. L.; Long, H. M.; Stylianou, J.; Thomas, W. A.; Leese, A.; Bell, A. I.; Bornkamm, G. W.; Mautner, J.; Rickinson, A. B.; Rowe, M. (2009). Jung, Jae U (ed.). "An Epstein-Barr Virus Anti-Apoptotic Protein Constitutively Expressed in Transformed Cells and Implicated in Burkitt Lymphomagenesis: The Wp/BHRF1 Link". PLoS Pathogens. 5 (3): e1000341. doi:10.1371/journal.ppat.1000341. PMC 2652661. PMID 19283066.
- Conrad NK, Fok V, Cazalla D, Borah S, Steitz JA (2006). "The challenge of viral snRNPs". Cold Spring Harb. Symp. Quant. Biol. 71: 377–384. doi:10.1101/sqb.2006.71.057. PMID 17381320. Retrieved 2010-08-13.
- Toczyski DP, Matera AG, Ward DC, Steitz JA (April 1994). "The Epstein-Barr virus (EBV) small RNA EBER1 binds and relocalizes ribosomal protein L22 in EBV-infected human B lymphocytes". Proc. Natl. Acad. Sci. U.S.A. 91 (8): 3463–3467. doi:10.1073/pnas.91.8.3463. PMC 43597. PMID 8159770. Retrieved 2010-08-13.