Death-associated protein 6
Death-associated protein 6 also known as Daxx is a protein that in humans is encoded by the DAXX gene.[5][6]



Function
Daxx, a Death domain-associated protein, was first discovered through its cytoplasmic interaction with the classical death receptor Fas. It has been associated with heterochromatin and PML-NBs (Promyelocytic Leukaemia nuclear bodies) and has been implicated in many nuclear processes including transcription and cell cycle regulation.
This gene encodes a multifunctional protein that resides in multiple locations in the nucleus and in the cytoplasm. Daxx serves as an H3.3 specific histone chaperone, interacting with an H3.3/H4 dimer.[7] It interacts with a wide variety of proteins, such as apoptosis antigen Fas, centromere protein C, and transcription factor erythroblastosis virus E26 oncogene homolog 1 (ETS1). In the nucleus, the encoded protein functions as a potent transcription repressor that binds to sumoylated transcription factors. Its repression can be relieved by the sequestration of this protein into promyelocytic leukemia nuclear bodies or nucleoli. This protein also associates with centromeres in G2 phase. In the cytoplasm, the encoded protein may function to regulate apoptosis. The subcellular localization and function of this protein are modulated by post-translational modifications, including sumoylation, phosphorylation and polyubiquitination.[8]
Structure and localization
Daxx is uniformly expressed throughout the body, except in the testes and thymus, which have especially high expression of the protein. At the level of the cell, Daxx is found in the cytoplasm, interacting with Fas-receptor or other cytoplasmic molecules, as well as in the nucleus, where it is interacting with some subnuclear structures. Several additional interacting proteins are known, but not always is there an understanding of the specific function and relevance of this interaction.
Nuclear
When the PML-NB are absent or disrupted, Daxx is delocalized and apoptosis does not occur. This interaction was demonstrated when PML-NB disrupted cells were treated and Daxx relocalized with the PML-NB. ATRX, a centromeric heterochromatin component co-localizes with Daxx. This partnership is found mainly in the S-phase of the cell cycle. No expression of Daxx leads to malfunction of S phase and cells with two nuclei are formed. Another centromeric component, CENP-C, associates with Daxx during interphase. While at first Daxx was said to be a “death protein”, it is suggested that associating with centromeric components leads to another function of Daxx.
Cytoplasmic and membrane
Fas-receptor stimulation causes Daxx to translocate out of the nucleus and into the cytoplasm. The breakdown of glucose produces reactive oxygen species (ROS). These induce extracellular Daxx to translocalize into the cytoplasm following an association with ASK1 (Apoptosis signal-regulating kinase1). Another mechanism for exogenous Daxx import involves CRM1. This transport mechanism is phosphorylation dependent. Nevertheless, it is not known whether the Fas-receptor stimuli or the ASK1 overexpression are caused by ROS or CRM1 mediated export.
Role in apoptosis
Fas-induced
After Fas stimulation, Daxx is activated and plays its role of pro-apoptotic protein in activating the c-JUN-N-Terminal Kinase (JNK) pathway. This pathway normally regulates stress-induced cell death. It is also essential for development of nerval system by programmed cell death. The real apoptotic process starts after activating this pathway. Daxx does not activate JNK itself but rather the upstream JNK kinase kinase ASK1. Some kind of positive feedback system was also discovered; JNK activates HIPK2, which stands for the translocation of nuclear Daxx to the cytoplasm. In turn, Daxx activates ASK1.
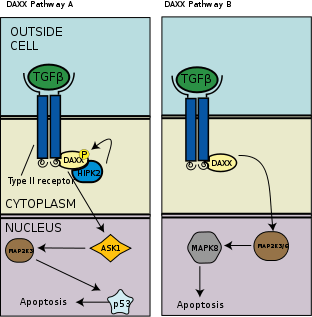
TGF-β regulated
TGF-β regulates a variety of different cellular developmental processes including growth, differentiation, proliferation, and cell death. Daxx interacts with the TGF-β type II receptor by binding of C-terminal domain of the protein. When the cell is treated with TGF-β, HIPK2, a nuclear kinase, phosphorylates Daxx and the activated Daxx in turn activates the JNK pathway (see "The Daxx Pathway" figure).
Miscellaneous mechanisms
Glucose breakdown produces ROS, which leads to Daxx production and relocalization, activating JNK pathway in turn. Another inducer of Daxx production is the exposure to UV-radiation. ASK1 will be transported to the nucleus when UV-irradiation is used to treat the cell. It is still unknown as to whether ASK1 binds Daxx, due to UV-irradiation. Another important cell death-property of Daxx is the association with PML-NB. It was shown that Daxx associates with Pml only when exposed to high oxidative stress or UV-irradiation. Another study showed loss of Daxx pro-apoptotic function in case of a mutant without Pml.
Anti-apoptotic function
A rather surprising property of Daxx is its anti-apoptotic function. When Daxx was not expressed or disrupted during embryonic development, it resulted in an early stage lethality. Other studies showed that lack of Daxx gene caused a higher apoptotic rate in embryonic stem cells. Only when Daxx was bound to Pml were apoptosis rates higher, suggesting that associated cytoplasmic Daxx has the role of an anti-apoptotic molecule.
Other functions
The omnipresence of Daxx in the cell nucleus suggests that the protein may also function as a transcription factor. Although it contains no known DNA-binding domains, Daxx can interact and suppress several transcription factors, such as p53, p73, and NF-κB. Proteins other than transcription factors are also blocked or inhibited by Daxx, such as the TGF-β pathway regulator, Smad4, conferring upon Daxx a major role in TGF-β signaling.
References
- ENSG00000206206, ENSG00000227046, ENSG00000206279, ENSG00000204209, ENSG00000229396 GRCh38: Ensembl release 89: ENSG00000231617, ENSG00000206206, ENSG00000227046, ENSG00000206279, ENSG00000204209, ENSG00000229396 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000002307 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Kiriakidou M, Driscoll DA, Lopez-Guisa JM, Strauss JF (1997). "Cloning and expression of primate Daxx cDNAs and mapping of the human gene to chromosome 6p21.3 in the MHC region". DNA Cell Biol. 16 (11): 1289–98. doi:10.1089/dna.1997.16.1289. PMID 9407001.
- Yang X, Khosravi-Far R, Chang HY, Baltimore D (1997). "Daxx, a novel Fas-binding protein that activates JNK and apoptosis". Cell. 89 (7): 1067–76. doi:10.1016/S0092-8674(00)80294-9. PMC 2989411. PMID 9215629.
- Lewis PW, Elsaesser SJ, Noh KM, Stadler SC, Allis CD (2010). "Daxx is an H3.3-specific histone chaperone and cooperates with ATRX in replication-independent chromatin assembly at telomeres". Proc. Natl. Acad. Sci. U.S.A. 107 (32): 140765–14080. doi:10.1073/pnas.1008850107. PMC 2922592. PMID 20651253.
- "Entrez Gene: DAXX".
Further reading
- Salomoni P, Khelifi AF (2006). "Daxx: death or survival protein?". Trends Cell Biol. 16 (2): 97–104. doi:10.1016/j.tcb.2005.12.002. PMID 16406523.
- Yang X, Khosravi-Far R, Chang HY, Baltimore D (1997). "Daxx, a novel Fas-binding protein that activates JNK and apoptosis". Cell. 89 (7): 1067–76. doi:10.1016/S0092-8674(00)80294-9. PMC 2989411. PMID 9215629.
- Suzuki Y, Yoshitomo-Nakagawa K, Maruyama K, Suyama A, Sugano S (1997). "Construction and characterization of a full length-enriched and a 5'-end-enriched cDNA library". Gene. 200 (1–2): 149–56. doi:10.1016/S0378-1119(97)00411-3. PMID 9373149.
- Ishov AM, Sotnikov AG, Negorev D, Vladimirova OV, Neff N, Kamitani T, Yeh ET, Strauss JF, Maul GG (1999). "PML is critical for ND10 formation and recruits the PML-interacting protein daxx to this nuclear structure when modified by SUMO-1". J. Cell Biol. 147 (2): 221–34. doi:10.1083/jcb.147.2.221. PMC 2174231. PMID 10525530.
- Li H, Leo C, Zhu J, Wu X, O'Neil J, Park EJ, Chen JD (2000). "Sequestration and inhibition of Daxx-mediated transcriptional repression by PML". Mol. Cell. Biol. 20 (5): 1784–96. doi:10.1128/MCB.20.5.1784-1796.2000. PMC 85360. PMID 10669754.
- Zhong S, Salomoni P, Ronchetti S, Guo A, Ruggero D, Pandolfi PP (2000). "Promyelocytic leukemia protein (PML) and Daxx participate in a novel nuclear pathway for apoptosis". J. Exp. Med. 191 (4): 631–40. doi:10.1084/jem.191.4.631. PMC 2195846. PMID 10684855.
- Li R, Pei H, Watson DK, Papas TS (2000). "EAP1/Daxx interacts with ETS1 and represses transcriptional activation of ETS1 target genes". Oncogene. 19 (6): 745–53. doi:10.1038/sj.onc.1203385. PMID 10698492.
- Charette SJ, Lavoie JN, Lambert H, Landry J (2000). "Inhibition of Daxx-mediated apoptosis by heat shock protein 27". Mol. Cell. Biol. 20 (20): 7602–12. doi:10.1128/MCB.20.20.7602-7612.2000. PMC 86317. PMID 11003656.
- Perlman R, Schiemann WP, Brooks MW, Lodish HF, Weinberg RA (2001). "TGF-beta-induced apoptosis is mediated by the adapter protein Daxx that facilitates JNK activation". Nat. Cell Biol. 3 (8): 708–14. doi:10.1038/35087019. PMID 11483955.
- Morozov VM, Gavrilova EV, Ogryzko VV, Ishov AM (2012). "Dualistic function of Daxx at centromeric and pericentromeric heterochromatin in normal and stress conditions". Nucleus. 3 (3): 276–85. doi:10.4161/nucl.20180. PMC 3414404. PMID 22572957.
External links
- DAXX+protein,+human at the US National Library of Medicine Medical Subject Headings (MeSH)
- PDBe-KB provides an overview of all the structure information available in the PDB for Human Death domain-associated protein 6
This article incorporates text from the United States National Library of Medicine, which is in the public domain.