ATX-II
ATX-II, also known as neurotoxin 2, Av2, Anemonia viridis toxin 2 or δ-AITX-Avd1c, is a neurotoxin derived from the venom of the sea anemone Anemonia sulcata. ATX-II slows down the inactivation of different voltage-gated sodium channels, including Nav1.1 and Nav1.2, thus prolonging action potentials.
| Molecular formula | C213 H329 N63 O61 S6 |
|---|---|
| Number of atoms | 672 |
| Molecular mass | 4940.70 Da |
| pI | 8.34 |
| Amino acid sequence | GVPCLCDSDGPSVRGNTLSGIIWLAGCPSGWHNCKKHGPTIGWCCKQ |
| Modifications | Disulfide bonds: 4:44, 6-34, 27-45 |
Sources
ATX-II is the main component of the venom of Mediterranean snakelocks sea anemone, Anemonia sulcata.[1][2] ATX-II is produced by the nematocysts in the sea anemone's tentacles and the anemone uses this venom to paralyze its prey.[3]
Etymology
"ATX-II" is an acronym for "anemone toxin".
Chemistry
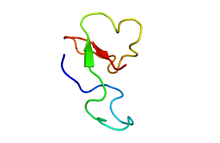
Structure
ATX-II is a protein comprising 47 amino acids[4] crosslinked by three disulfide bridges. The molecular mass of the protein is 4,94 kDa (calculated with ProtParam ExPASy).
Family and homology
ATX-II belongs to the sea anemone neurotoxin family. Purification studies of ATX-II and the two other sea anemone neurotoxins, I and III, have revealed the polypeptide nature of these toxins.[5] Toxins I and II are very potent paralyzing toxins that act on crustaceans, fish and mammals and have cardiotoxic and neurotoxic effects.[6] Toxin III has been shown to cause muscular contraction with subsequent paralysis in the crab Carcinus maenas. All three toxins are highly homologous and block neuromuscular transmission in crabs.[3]
Four other sea anemone toxins purified from Condylactis aurantiaca show close sequence similarities with toxins I, II and III of Anemonia sulcata.[7] The effect of these different toxins on Carcinus meanas is visually indistinguishable, namely cramp followed by paralysis and death. However, their mode of action differs. Toxin IV of Condylactis aurantiaca causes a repetitive firing of the excitatory axon for several minutes resulting in muscle contraction without causing a detectable change in the amplitude of the excitatory junction potentials (EJPS). In contrast, the application of Toxin II from Anemonia sulcata results in the increase of the EJPS up to 40 mV causing large action potentials at the muscle fibers.[8] Other toxins with a similar mode of action to ATX-II are α-scorpion toxins. Although both sea anemone and α-scorpion toxins bind to common overlapping elements on the extracellular surface of sodium channels, they belong to distinct families and share no sequence homology.[9] The toxins AFT-II (from Anthopleura fuscoviridis) and ATX-II differ by only one amino acid, L36A, and the protein sequence of BcIII (from Bunodosoma caissarum) is 70% similar to ATX-II.[10]
Target
ATX-II is highly potent at voltage-gated sodium channels subtype 1.1 and 1.2 (Nav1.1 and Nav1.2) with an EC50 of approximately 7 nM when tested in human embryonic kidney 293 cells lines.[11] Moreover, studies suggest that ATX-II interacts with glutamic acid residue (Glu-1613 and 1616 in Nav1.2) on the third and fourth transmembrane loop (S3-S4) of domain IV on the alpha-subunit of neuronal channel Nav1.2 in rats.[12]
The KD of type IIa Na+ channels for ATX II is 76 ± 6 nM. In small and large dorsal root ganglion cells mainly Nav1.1, Nav1.2 and Nav1.6 are sensitive to ATX-II.[13] The binding of the toxin can only occur when the sodium channel is open.[14]
Mode of action
The major action of ATX-II is to delay sodium channel inactivation. Studies using giant crayfish axons and myelinated fibers from frogs indicate that ATX-II acts at low doses, without changing the opening mechanism or steady-state potassium conductance.[14] This mode of action is caused by binding of ATX-II across the extracellular loop. ATX-II slows conformational changes or translocation that are necessary for closing the sodium channel. When applied externally in high concentrations (100 μM range), ATX-II reduces potassium conductance, yet without modifying the kinetic properties of the potassium channel.[15]
ATX-II prolongs the duration of the cardiac action potential, as demonstrated in cultured embryonic chicken cardiac muscle cells.[16] ATX-II also selectively activates A-fibers of peripheral nerves projecting to the sensory neuron of the dorsal root ganglia (DRG) by enhancing resurging currents in DRGs. This mechanism can thereby induce itch-like sensations and pain.[13]
Toxicity
People who got in contact with Anemonia sulcata reported symptoms such as pain and itch. The same symptoms were found in human research subjects after injection of ATX-II into their skin.[13]
In cardiac muscle tissue of various mammals, ATX-II has been shown to produce large and potentially lethal increases in heart rate.[17] The lethal dose of ATX-II for the crab Carcinus maenas is 2 μg/kg.[5]
References
- UniProt. Taxonomy - Anemonia sulcata (Mediterranean snakelocks sea anemone) [Internet]. UniProt. [cited 2017Oct2]. Available from: https://www.uniprot.org/taxonomy/6108
- UniProt. Delta-actitoxin-Avd1c [Internet]. Uniprot. 2017 [cited 2017Oct2]. Available from: https://www.uniprot.org/uniprot/P01528.
- Béress L. Isolation and characterisation of three polypeptides with neurotoxic activity from Anemonia sulcata. FEBS Letters. 1975;50(3):311–4.
- Wunderer G, Fritz H, Wachter E, Machleidt W. Amino-Acid Sequence of a Coelenterate Toxin: Toxin II from Anemonia sulcata. European Journal of Biochemistry. 1976;68(1):193–8.
- Béress L, Béress R, Wunderer G. Purification of three polypeptides with neuroand cardiotoxic activity from the sea anemone Anemonia sulcata. Toxicon. 1975;13(5):359–64.
- Beress, L., and R. Beress. "Reinigung zweier krabbenlähmender Toxine aus der Seeanemone Anemonia sulcata." Kieler Meeresforsch 27 (1971): 117-127.
- Béress R, Wunderer G, Béress L. Neurotoxins from Sea Anemones. Purification and Characterisation of Four Polypeptides with Neurotoxic Activity fromCondylactis aurantiaca. Hoppe-Seyler's Zeitschrift für physiologische Chemie. 1976;357(1):409–14.
- Rathmayer, W., Barbara Jessen, and L. Beress. "Effect of toxins of sea anemones on neuromuscular transmission." Naturwissenschaften 62.11 (1975): 538-539.
- Catterall WA. Neurotoxins that Act on Voltage-Sensitive Sodium Channels in Excitable Membranes. Annual Review of Pharmacology and Toxicology. 1980;20(1):15–43.
- Oliveira JS, Redaelli E, Zaharenko AJ, Cassulini RR, Konno K, Pimenta DC, et al. Binding Specificity of Sea Anemone Toxins to Nav1.1-1.6 Sodium Channels. Journal of Biological Chemistry. 2004;279(32):33323–35.
- Spencer CI. Actions of ATX-II and other gating-modifiers on Na(+) currents in HEK-293 cells expressing WT and DeltaKPQ hNa(V) 1.5 Na(+) channels. Toxicon. 2009 Jan;53(1):78-89.
- Rogers JC, Qu Y, Tanada TN, Scheuer T, Catterall WA. Molecular Determinants of High Affinity Binding of Alpha-Scorpion Toxin and Sea Anemone Toxin in the S3-S4 Extracellular Loop in Domain IV of the Na Channel Alpha-Subunit. Journal of Biological Chemistry. 1996May;271(27):15950–62.
- Klinger AB, Eberhardt M, Link AS, Namer B, Kutsche LK, Schuy ET, et al. Sea-Anemone Toxin ATX-II Elicits A-Fiber-Dependent Pain and Enhances Resurgent and Persistent Sodium Currents in Large Sensory Neurons. Molecular Pain. 2012;8.
- Romey G, Abita JP, Schweitz H, Wunderer G, Lazdunski. Sea anemone toxin:a tool to study molecular mechanisms of nerve conduction and excitation-secretion coupling. Proceedings of the National Academy of Sciences. 1976Jan;73(11):4055–9.
- Bergman C, Dubois J, Rojas E, Rathmayer W. Decreased rate of sodium conductance inactivation in the node of Ranvier induced by a polypeptide toxin from sea anemone. Biochimica et Biophysica Acta (BBA) - Biomembranes. 1976;455(1):173–84.
- Romey, G., et al. "Pharmacological properties of the interaction of a sea anemone polypeptide toxin with cardiac cells in culture." Journal of Pharmacology and Experimental Therapeutics 213.3 (1980): 607-615.
- Alsen C. Biological significance of peptides from Anemonia sulcata. Fed Proc. 1983;42(1):101-8.