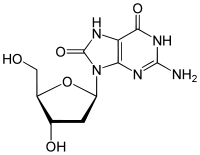
8-Oxo-2'-deoxyguanosine
8-Oxo-2'-deoxyguanosine (8-oxo-dG) is an oxidized derivative of deoxyguanosine. 8-Oxo-dG is one of the major products of DNA oxidation.[1] Concentrations of 8-oxo-dG within a cell are a measurement of oxidative stress.
 | |
| Names | |
|---|---|
| IUPAC name
2-amino-9-[(2R,4S,5R)-4-hydroxy-5-(hydroxymethyl)oxolan-2-yl]-3,7-dihydropurine-6,8-dione | |
| Other names
7,8-Dihydro-8-oxo-2'-deoxyguanosine; 7,8-Dihydro-8-oxodeoxyguanosine; 8-Hydroxy-2'-deoxyguanosine; 8-Hydroxydeoxyguanosine; 8-Oxo-2'-deoxyguanosine; 8-Oxo-7,8-dihydro-2'-deoxyguanosine; 8-Oxo-7,8-dihydrodeoxyguanosine; 8-Oxo-dG; 8-OH-dG | |
| Identifiers | |
3D model (JSmol) |
|
| ChEBI | |
| ChemSpider | |
PubChem CID |
|
| UNII | |
CompTox Dashboard (EPA) |
|
| |
| |
| Properties | |
| C10H13N5O5 | |
| Molar mass | 283.24 g/mol |
Except where otherwise noted, data are given for materials in their standard state (at 25 °C [77 °F], 100 kPa). | |
| Infobox references | |
In DNA
_and_with_tumorigenesis_(B)._Brown_shows_8-oxo-dG.jpg)
Steady-state levels of DNA damages represent the balance between formation and repair. Swenberg et al.[3] measured average frequencies of steady state endogenous DNA damages in mammalian cells. The most frequent oxidative DNA damage normally present in DNA is 8-oxo-dG, occurring at an average frequency of 2,400 per cell.
When 8-oxo-dG is induced by a DNA damaging agent it is rapidly repaired. For example, 8-oxo-dG was increased 10-fold in the livers of mice subjected to ionizing radiation, but the excess 8-oxo-dG was rapidly removed with a half-life of 11 minutes.[4]
As reviewed by Valavanidis et al.[5] increased levels of 8-oxo-dG in a tissue can serve as a biomarker of oxidative stress. They also noted that increased levels of 8-oxo-dG are frequently found during carcinogenesis.
In the figure shown in this section, the colonic epithelium from a mouse on a normal diet has a low level of 8-oxo-dG in its colonic crypts (panel A). However, a mouse likely undergoing colonic tumorigenesis (due to deoxycholate added to its diet[2]) has a high level of 8-oxo-dG in its colonic epithelium (panel B). Deoxycholate increases intracellular production of reactive oxygen resulting in increased oxidative stress,[6][7] and this leads to tumorigenesis and carcinogenesis. Of 22 mice fed the diet supplemented with deoxycholate, 20 (91%) developed colonic tumors after 10 months on the diet, and the tumors in 10 of these mice (45% of mice) included an adenocarcinoma (cancer).[2]
In aging
8-oxo-dG increases with age in DNA of mammalian tissues.[8] 8-oxo-dG increases in both mitochondrial DNA and nuclear DNA with age.[9] Fraga et al.[10] estimated that in rat kidney, for every 54 residues of 8-oxo-dG repaired, one residue remains unrepaired. (See also DNA damage theory of aging.)
In carcinogenesis
Valavanidis et al.[5] pointed out that oxidative DNA damage, such as 8-oxo-dG, likely contributes to carcinogenesis by two mechanisms. The first mechanism involves modulation of gene expression, whereas the second is through the induction of mutations.
Epigenetic alterations
Epigenetic alteration, for instance by methylation of CpG islands in a promoter region of a gene, can repress expression of the gene (see DNA methylation). In general, epigenetic alteration can modulate gene expression. As reviewed by Bernstein and Bernstein,[11] the repair of various types of DNA damages can, with low frequency, leave remnants of the different repair processes and thereby cause epigenetic alterations. 8-Oxo-dG is primarily repaired by base excision repair (BER).[12] Li et al.[13] reviewed studies indicating that one or more BER proteins also participate(s) in epigenetic alterations involving DNA methylation, demethylation or reactions coupled to histone modification. Nishida et al.[14] examined 8-oxo-dG levels and also evaluated promoter methylation of 11 tumor suppressor genes (TSGs) in 128 liver biopsy samples. These biopsies were taken from patients with chronic hepatitis C, a condition causing oxidative damages in the liver. Among 5 factors evaluated, only increased levels of 8-oxo-dG was highly correlated with promoter methylation of TSGs (p<0.0001). This promoter methylation could have reduced expression of these tumor suppressor genes and contributed to carcinogenesis.
Mutagenesis
Yasui et al.[15] examined the fate of 8-oxo-dG when this oxidized derivative of deoxyguanosine was inserted into the thymidine kinase gene in a chromosome within human lymphoblastoid cells in culture. They inserted 8-oxo-dG into about 800 cells, and could detect the products that occurred after the insertion of this altered base, as determined from the clones produced after growth of the cells. 8-Oxo-dG was restored to G in 86% of the clones, probably reflecting accurate base excision repair or translesion synthesis without mutation. G:C to T:A transversions occurred in 5.9% of the clones, single base deletions in 2.1% and G:C to C:G transversions in 1.2%. Together, these more common mutations totaled 9.2% of the 14% of mutations generated at the site of the 8-oxo-dG insertion. Among the other mutations in the 800 clones analyzed, there were also 3 larger deletions, of sizes 6, 33 and 135 base pairs. Thus 8-oxo-dG, if not repaired, can directly cause frequent mutations, some of which may contribute to carcinogenesis.
In memory formation
Two reviews[16][17] summarize the large body of evidence, reported largely between 1996 and 2011, for the critical and essential role of ROS in memory formation. A recent additional body of evidence indicates that both the formation and storage of memory depend on epigenetic modifications in neurons, including alterations in neuronal DNA methylation.[18][19] The two bodies of information on memory formation appear to be connected in 2016 by the work of Zhou et al,[20] who showed that 8-oxo-dG, a major product of ROS interaction with DNA,[21][22] has a central role in epigenetic DNA demethylation.
The activation of transcription of some genes by transcription factors depends on the presence of 8-oxo-dG in the promoter regions and its recognition by the DNA repair glycosylase OGG1.[23][22]
As reviewed by Duke et al., neuron DNA methylation and demethylation are altered by neuronal activity. Active DNA methylations and demethylations are required for synaptic plasticity, are modified by experiences, and are required for memory formation and maintenance.[24]
In mammals, DNA methyltransferases (which add methyl groups to DNA bases) exhibit a strong sequence preference for cytosines within the particular DNA sequence cytosine-phosphate-guanine (CpG sites).[25] In the mouse brain, 4.2% of all cytosines are methylated, primarily in the context of CpG sites, forming 5mCpG.[26] Most hypermethylated 5mCpG sites increase the repression of associated genes.[26] As shown by Zhou et al.,[20] and illustrated below, oxidation of the guanine in the methylated CpG site, to form 5mCp-8-oxo-dG is the first step in demethylation.
8-oxo-dG complexed with OGG1 likely has a major role in facilitating thousands of rapid demethylations of methylated cytosines in CpG sites during formation of memory and further demethylations (over a period of weeks) during memory consolidation. As shown in 2016 by Halder et al.[27] using mice, and in 2017 by Duke et al.[24] using rats, when contextual fear conditioning is applied to the rodents, causing an especially strong long-term memory to form, within hours there are thousands of methylations and demethylations in the hippocampus brain region neurons. As shown with the rats, 9.2% of the genes in the rat hippocampus neurons are differentially methylated. In mice, examined at 4 weeks after conditioning, the hippocampus methylations and demethylations were reversed (the hippocampus is needed to form memories but memories are not stored there) while substantial differential CpG methylation and demethylation occurred in cortical neurons during memory maintenance. There were 1,223 differentially methylated genes in the anterior cingulate cortex of mice four weeks after contextual fear conditioning. Where demethylations occur, oxidation of the guanine in the CpG site to form 8-oxo-dG is an important first step.[20]
Demethylation at CpG sites requires 8-oxo-dG

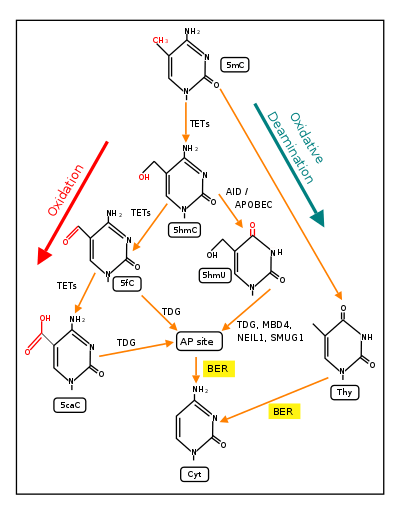
TET1 is a key enzyme involved in demethylating 5mCpG. However, TET1 is only able to act on 5mCpG if an ROS has first acted on the guanine to form 8-hydroxy-2'-deoxyguanosine (8-OHdG or its tautomer 8-oxo-dG), resulting in a 5mCp-8-OHdG dinucleotide (see first figure in this section).[20] After formation of 5mCp-8-OHdG, the base excision repair enzyme OGG1 binds to the 8-OHdG lesion without immediate excision. Adherence of OGG1 to the 5mCp-8-OHdG site recruits TET1, allowing TET1 to oxidize the 5mC adjacent to 8-OHdG, as shown in the first figure in this section. This initiates the demethylation pathway shown in the second figure in this section.
Altered protein expression in neurons, controlled by 8-oxo-dG-dependent demethylation of CpG sites in gene promoters within neuron DNA, is central to memory formation.[29]
See also
References
- Nadja C. de Souza-Pinto; Lars Eide; Barbara A. Hogue; Tanja Thybo; Tinna Stevnsner; Erling Seeberg; Arne Klungland & Vilhelm A. Bohr (July 2001). "Repair of 8-Oxodeoxyguanosine Lesions in Mitochondrial DNA Depends on the Oxoguanine DNA Glycosylase (OGG1) Gene and 8-Oxoguanine Accumulates in the Mitochondrial DNA of OGG1-defective Mice". Cancer Research. 61 (14): 5378–5381. PMID 11454679.
- Prasad AR, Prasad S, Nguyen H, Facista A, Lewis C, Zaitlin B, Bernstein H, Bernstein C (2014). "Novel diet-related mouse model of colon cancer parallels human colon cancer". World J Gastrointest Oncol. 6 (7): 225–43. doi:10.4251/wjgo.v6.i7.225. PMC 4092339. PMID 25024814.
- Swenberg, J. A.; Lu, K.; Moeller, B. C.; Gao, L.; Upton, P. B.; Nakamura, J.; Starr, T. B. (2011). "Endogenous versus Exogenous DNA Adducts: Their Role in Carcinogenesis, Epidemiology, and Risk Assessment". Toxicological Sciences. 120: S130–S145. doi:10.1093/toxsci/kfq371. PMC 3043087. PMID 21163908.
- Hamilton ML, Guo Z, Fuller CD, Van Remmen H, Ward WF, Austad SN, Troyer DA, Thompson I, Richardson A (2001). "A reliable assessment of 8-oxo-2-deoxyguanosine levels in nuclear and mitochondrial DNA using the sodium iodide method to isolate DNA". Nucleic Acids Res. 29 (10): 2117–26. doi:10.1093/nar/29.10.2117. PMC 55450. PMID 11353081.
- Valavanidis A, Vlachogianni T, Fiotakis K, Loridas S (2013). "Pulmonary oxidative stress, inflammation and cancer: respirable particulate matter, fibrous dusts and ozone as major causes of lung carcinogenesis through reactive oxygen species mechanisms". Int J Environ Res Public Health. 10 (9): 3886–907. doi:10.3390/ijerph10093886. PMC 3799517. PMID 23985773.
- Tsuei, Jessica; Chau, Thinh; Mills, David; Wan, Yu-Jui Yvonne (2014). "Bile acid dysregulation, gut dysbiosis, and gastrointestinal cancer". Experimental Biology and Medicine. 239 (11): 1489–1504. doi:10.1177/1535370214538743. PMC 4357421. PMID 24951470.
- Ajouz, Hana; Mukherji, Deborah; Shamseddine, Ali (2014). "Secondary bile acids: An underrecognized cause of colon cancer". World Journal of Surgical Oncology. 12: 164. doi:10.1186/1477-7819-12-164. PMC 4041630. PMID 24884764.
- Nie B, Gan W, Shi F, Hu GX, Chen LG, Hayakawa H, Sekiguchi M, Cai JP (2013). "Age-dependent accumulation of 8-oxoguanine in the DNA and RNA in various rat tissues". Oxid Med Cell Longev. 2013: 303181. doi:10.1155/2013/303181. PMC 3657452. PMID 23738036.
- Hamilton ML, Van Remmen H, Drake JA, Yang H, Guo ZM, Kewitt K, Walter CA, Richardson A (2001). "Does oxidative damage to DNA increase with age?". Proc. Natl. Acad. Sci. U.S.A. 98 (18): 10469–74. Bibcode:2001PNAS...9810469H. doi:10.1073/pnas.171202698. PMC 56984. PMID 11517304.
- Fraga CG, Shigenaga MK, Park JW, Degan P, Ames BN (1990). "Oxidative damage to DNA during aging: 8-hydroxy-2'-deoxyguanosine in rat organ DNA and urine". Proc. Natl. Acad. Sci. U.S.A. 87 (12): 4533–7. Bibcode:1990PNAS...87.4533F. doi:10.1073/pnas.87.12.4533. PMC 54150. PMID 2352934.
- Bernstein C, Bernstein H (2015). "Epigenetic reduction of DNA repair in progression to gastrointestinal cancer". World J Gastrointest Oncol. 7 (5): 30–46. doi:10.4251/wjgo.v7.i5.30. PMC 4434036. PMID 25987950.
- Scott TL, Rangaswamy S, Wicker CA, Izumi T (2014). "Repair of oxidative DNA damage and cancer: recent progress in DNA base excision repair". Antioxid. Redox Signal. 20 (4): 708–26. doi:10.1089/ars.2013.5529. PMC 3960848. PMID 23901781.
- Li J, Braganza A, Sobol RW (2013). "Base excision repair facilitates a functional relationship between Guanine oxidation and histone demethylation". Antioxid. Redox Signal. 18 (18): 2429–43. doi:10.1089/ars.2012.5107. PMC 3671628. PMID 23311711.
- Nishida N, Arizumi T, Takita M, Kitai S, Yada N, Hagiwara S, Inoue T, Minami Y, Ueshima K, Sakurai T, Kudo M (2013). "Reactive oxygen species induce epigenetic instability through the formation of 8-hydroxydeoxyguanosine in human hepatocarcinogenesis". Dig Dis. 31 (5–6): 459–66. doi:10.1159/000355245. PMID 24281021.
- Yasui M, Kanemaru Y, Kamoshita N, Suzuki T, Arakawa T, Honma M (2014). "Tracing the fates of site-specifically introduced DNA adducts in the human genome". DNA Repair (Amst.). 15: 11–20. doi:10.1016/j.dnarep.2014.01.003. PMID 24559511.
- Massaad CA, Klann E (May 2011). "Reactive oxygen species in the regulation of synaptic plasticity and memory". Antioxid. Redox Signal. 14 (10): 2013–54. doi:10.1089/ars.2010.3208. PMC 3078504. PMID 20649473.
- Beckhauser TF, Francis-Oliveira J, De Pasquale R (2016). "Reactive Oxygen Species: Physiological and Physiopathological Effects on Synaptic Plasticity". J Exp Neurosci. 10 (Suppl 1): 23–48. doi:10.4137/JEN.S39887. PMC 5012454. PMID 27625575.
- Day JJ, Sweatt JD (January 2011). "Epigenetic modifications in neurons are essential for formation and storage of behavioral memory". Neuropsychopharmacology. 36 (1): 357–8. doi:10.1038/npp.2010.125. PMC 3055499. PMID 21116250.
- Sweatt JD (October 2016). "Neural plasticity and behavior - sixty years of conceptual advances". J. Neurochem. 139 Suppl 2: 179–199. doi:10.1111/jnc.13580. PMID 26875778.
- Zhou X, Zhuang Z, Wang W, He L, Wu H, Cao Y, Pan F, Zhao J, Hu Z, Sekhar C, Guo Z (September 2016). "OGG1 is essential in oxidative stress induced DNA demethylation". Cell. Signal. 28 (9): 1163–71. doi:10.1016/j.cellsig.2016.05.021. PMID 27251462.
- Jena NR (July 2012). "DNA damage by reactive species: Mechanisms, mutation and repair". J. Biosci. 37 (3): 503–17. doi:10.1007/s12038-012-9218-2. PMID 22750987.
- Ba X, Boldogh I (April 2018). "8-Oxoguanine DNA glycosylase 1: Beyond repair of the oxidatively modified base lesions". Redox Biol. 14: 669–678. doi:10.1016/j.redox.2017.11.008. PMC 5975208. PMID 29175754.
- Seifermann M, Epe B (June 2017). "Oxidatively generated base modifications in DNA: Not only carcinogenic risk factor but also regulatory mark?". Free Radic. Biol. Med. 107: 258–265. doi:10.1016/j.freeradbiomed.2016.11.018. PMID 27871818.
- Duke CG, Kennedy AJ, Gavin CF, Day JJ, Sweatt JD (July 2017). "Experience-dependent epigenomic reorganization in the hippocampus". Learn. Mem. 24 (7): 278–288. doi:10.1101/lm.045112.117. PMC 5473107. PMID 28620075.
- Ziller MJ, Müller F, Liao J, Zhang Y, Gu H, Bock C, Boyle P, Epstein CB, Bernstein BE, Lengauer T, Gnirke A, Meissner A (December 2011). "Genomic distribution and inter-sample variation of non-CpG methylation across human cell types". PLoS Genet. 7 (12): e1002389. doi:10.1371/journal.pgen.1002389. PMC 3234221. PMID 22174693.
- Fasolino M, Zhou Z (May 2017). "The Crucial Role of DNA Methylation and MeCP2 in Neuronal Function". Genes (Basel). 8 (5): 141. doi:10.3390/genes8050141. PMC 5448015. PMID 28505093.
- Halder R, Hennion M, Vidal RO, Shomroni O, Rahman RU, Rajput A, Centeno TP, van Bebber F, Capece V, Garcia Vizcaino JC, Schuetz AL, Burkhardt S, Benito E, Navarro Sala M, Javan SB, Haass C, Schmid B, Fischer A, Bonn S (January 2016). "DNA methylation changes in plasticity genes accompany the formation and maintenance of memory". Nat. Neurosci. 19 (1): 102–10. doi:10.1038/nn.4194. PMC 4700510. PMID 26656643.
- Bayraktar G, Kreutz MR (2018). "The Role of Activity-Dependent DNA Demethylation in the Adult Brain and in Neurological Disorders". Front Mol Neurosci. 11: 169. doi:10.3389/fnmol.2018.00169. PMC 5975432. PMID 29875631.
- Day JJ, Sweatt JD (November 2010). "DNA methylation and memory formation". Nat. Neurosci. 13 (11): 1319–23. doi:10.1038/nn.2666. PMC 3130618. PMID 20975755.