The Ant and the Peacock
"The Ant and the Peacock" is a quip formulated to express a perceived problem in evolutionary theory during the first half of the twentieth century. The problems were altruism as epitomized by the ant and other social insects, and potentially debilitating display signals such as the peacock's tail. These problems have an interesting history in evolutionary biology and highlight more the power of social and political forces in shaping science than any honest difficulty with the problems. The solution of both of these problems were made popular in the 1975 with E. O. Wilson's publication of sociobiology.[1] His ideas would later be used to define and direct the work of researchers in evolutionary psychology.
| We're all Homo here Evolution |
| Relevant Hominids |
| A Gradual Science |
| Plain Monkey Business |
v - t - e |
The problems
The ant and peacock represent problems because natural selection cannot select for a trait which is overall more costly to the given organism than any benefit it receives. This is a hallmark of the theory, and any derivations from it could throw the whole issue into serious doubt.
The ant represents the problem of altruism, which is helping another member of your species at a cost to yourself. The peacock's tail is different trait, but the same essential problem. The tail is very costly to the male peacock both to produce and to have. It makes it more likely to be seen by predators and easier to catch. Both of these traits would seem to violate the central tenet of the theory of natural selection.
The solutions
The Ant
The problem of altruism was solved first, most likely because its solution was discoverable in mathematics as much as observation. At the heart of the idea is the concept of "inclusive fitness", developed by W. D. Hamilton, which points out that the fodder for selection of traits is not merely the direct benefits of that trait but any indirect benefits that are produced as well. While indirect benefits are less valuable than direct benefits, a trait that produces enough indirect benefits could still easily evolve if it has no direct benefits — or maybe even a cost to direct fitness.
This idea solves the ant problem in several different ways. The first is, through nepotism, benefits given to relatives that increase their likelihood to produce offspring are indirect benefits to the giver. Probability theory can be used to show that a given offspring of an organism will have about 50% of that organism's genes. The offspring of that organism's siblings will have about 25% of the organism's genes. So it should be fairly easy to see that helping your brothers or sisters produce two more offspring is evolutionarily equivalent to producing one direct offspring (from a gene's point of view). So an organism will take a cost to its direct fitness, if it increases the fitness of a relative in such a way that it is greater than the initial cost. This is represented by the equation 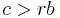 with c = cost, r = percent relatedness of a relative, and b equal to the benefit to that relative. This solves the problem of many social insects such as the ants, bees, and wasps, because the workers — who never produce offspring — are all at least half siblings, being the spawn of a single queen (although they may have different fathers). The workers — or their genes, which is what really matters — then receive enough indirect benefit from helping the queen produce more offspring that they do not need to have any of their own.
with c = cost, r = percent relatedness of a relative, and b equal to the benefit to that relative. This solves the problem of many social insects such as the ants, bees, and wasps, because the workers — who never produce offspring — are all at least half siblings, being the spawn of a single queen (although they may have different fathers). The workers — or their genes, which is what really matters — then receive enough indirect benefit from helping the queen produce more offspring that they do not need to have any of their own.
Indirect fitness
Indirect fitness also explains altruism between non-relatives. Since the giver cannot gain indirect benefit from offspring produced by the receiver, something else must be going on. Usually it is a form of reciprocal altruism - essentially an "I will scratch your back if you scratch my back" scenario. Because many problems encountered in the environment are describable as a non-zero-sum game, the absolute benefits to both parties can be greater than the actual costs. One can think about this by looking at hunting. Hunting is at best a crude mechanism for bringing in food. It is a "boom or bust" system: you either have a lot of meat or you have no meat at all. A successful hunt will produce more meat than you can possibly eat before it will rot and become worthless. By giving the portion of meat that is likely to rot before you can eat it to your neighbor who did not have a successful hunt, you incur little cost. Your neighbor, though, receives a massive benefit, and if that hunter then returns the favor the next time the situation is reversed, you too will receive a massive benefit. Both parties gain far more than the individual costs.
The Peacock
The solution of the peacock's tail problem is really to recognize that direct survival fitness is not the only possible selection force. Another force just as powerful, if not more so, is sexual selection, and at the heart of sexual selection is female choice. The minimum reproductive investment needed to produce successful offspring is very different between the males of most species and the females. Males usually only need to invest a few gamete cells and a few minutes of time, while females are required to gestate the offspring (or at least produce eggs, which require far more energy than sperm) for a certain length of time, give birth, and care for it till it is ready to be out on its own. The limiting factor in a male's reproductive success is the number of females he mates with; the limiting factor for a female is usually the quality of the male. This means that males can get away with not being very picky about who they are willing to mate with, but females must be very picky.
Males then are under intensive selection from females to signal quality and fitness. Signals of quality have to meet several requirements to work. The first is they have to honestly signal quality – if a trait is easy to fake it is worthless – and secondly, high quality versions of the trait must be preferred by the females of the species over low quality versions. One of the easiest ways to ensure that a signal is honest is to make it costly to produce and maintain. The more costly the signal is, the more honest it will have to be. Essentially this is the idea of handicapping, where a male with a trait that is very dangerous to produce and maintain, who still seems to be doing fine, is saying "wow, even when I make life more difficult for myself, I am still awesome." Indeed, experiments have shown that chicks fathered by peacocks with larger tails and more eye spots are healthier on hatching and more likely to survive. [2]
The effect from this quality signal is also self-perpetuating to a degree, in that the impressive tail is a heritable trait; if a female mates with a peacock that has an impressive tail, she is latching her own genes onto the same strand of DNA with the genes for impressive tails. Thus, if a female chooses an attractive mate, her own male offspring will reproduce more effectively.
The last part, female preference, can be almost any arbitrary signal. In the case of the peacock, it is a large brightly colored tail, but in other species it might be complexity of nest building, territory maintained, giant fatty deposits in the cheeks, or one of many other examples.
Social and political history
It is interesting to note that Charles Darwin actually published hypotheses about mechanisms that could have explained both traits. In particular, his writing on sexual selection in his later works offered a complete description of the solution of the peacock's tail. His work on this was largely ignored because Victorian England had a hard time accepting the concept of "female choice". The standard view of females being delicate, indecisive creatures in need of strong male guidance from birth till death was so ingrained that the thought that female choice could be one of the most significant selective forces in evolutionary theory seemed laughable. In this regard Alfred Russel Wallace, who was co-discoverer of the theory of evolution (due mostly to historical accident and Darwin's inherent reluctance to publish), won the debate, at least temporarily. Wallace, a good Christian patriarch, assured his readers that the peacock's tail would remain a mystery for a hundred years.
Female choice was ignored completely in evolutionary models until the 1950s, when consensus reluctantly agreed that there was some choice going on. But this was written off as only making sure that females mated with the same species. It was thought that without some obvious display signal, that made sure the females knew what was going on, they would likely mate with just about anything. It wasn't until after the 1960s and into the 1970s that researchers began to theorize in terms of actual female choice. Many then turned back to the original works of Darwin to see that the current "new" ideas had been there all along but ignored.
Female choice is now finally recognized as a powerful force in selection and evolution, but it is being attacked now by an interesting faction. Many of those in the more ultra-extreme divisions of the feminist movement see the description of female choice as denigrating to women. Many seem to think all of evolutionary history is purely a mechanism to repress women, and any description of women as a filter for display signals in males is inherently sexist. The struggle continues to this day, mostly with evolutionary psychologists, who attempt to use female choice, sexual selection, and other evolutionary mechanisms to specifically address issues in human psychology.
References
- Sociobiology: The New Synthesis, Twenty-Fifth Anniversary Edition by E. O. Wilson (2000) Belknap Press, 2nd ed. ISBN 0674002350.
- Tale of the Peacock