Recent African Origin hypothesis
The Recent African Origin[1] (RAO) hypothesis is a model of human evolution which argues that anatomically-modern humans had an exclusive origin in Africa within the last 200,000 years.[2] Migrations out of Africa (<120,000 BP), RAO alleges, replaced archaic humans (such as Neanderthals) across the Old World with negligible to no interbreeding.
| We're all Homo here Evolution |
| Relevant Hominids |
| A Gradual Science |
| Plain Monkey Business |
v - t - e |
Neanderthal-derived genes in living Eurasians are currently estimated to be 1.5 to 2.1%,[3] with additional Denisovan genes in Papua New Guineans and Australian aborigines. This small level of archaic admixture is compatible with the RAO: "a recent African origin (RAO) model for modern humans is still supported by the data"[4][5][6] (see Misconceptions below).
In 2015 researchers discovered that 7.4% of the genome of an early Upper Paleolithic modern human from Peştera cu Oase (Romania) was Neanderthal.[7] This higher level of genetic admixture has been explained by the extensive change in modern-human gene pools (through population-size changes and migrations) during the start of the Holocene, and by presuming that Neanderthal (and perhaps other archaic) contributions have been reduced.[8] Since archaic genetic contributions were formerly higher, some now argue that "Neandertal and Denisova genomic data are most commensurate with the Assimilation model"[9][10][11] (note though these higher admixture rates have been questioned as a possible overestimation[12] and one specimen whose genome was sequenced in 2014 - a femur from western Siberia, dated 45,000 BP, showed only a very small higher admixture percentage, 2.3%[13]).
Misconceptions
The RAO model is usually misinterpreted as denying any hybridization between African moderns and non-African archaics took place. However the RAO hypothesis has never (since it was formulated, e.g. Cann et al. 1987) ruled out the possibility there was minimal interbreeding.[14][15][16][17][18] Archaic genetic input though is "considered essentially insignificant" (and therefore negligible) which contrasts RAO to the Assimilation model (AM).[19] Brauer et al. (2004) criticize Multiregionalists for setting up a straw man argument that RAO strictly posits no interbreeding occurred, which is easy to refute (because the least possible amount of archaic admixture would then falsify the hypothesis):
Complete replacement does not feature in all versions of the Out of Africa hypothesis, and therefore we contend that it would not be falsified by demonstration of some modern-archaic human gene flow outside of Africa. Both before and after the pioneering study by Cann et al. (1987), proponents of the Out of Africa hypothesis accepted the possibility of a certain amount of gene flow between the migrating early anatomically modern humans and the non-African archaic groups (Brauer, 1984, 1992, 2001; Cann, 1992; Stringer, 1992, 2001; Stringer and Brauer, 1994).
Exactly who has ever claimed absolutely no interbreeding occurred is unclear (not even Rebecca Cann in her influential mtDNA studies rejected slight Neanderthal admixture). Nevertheless Stringer has called this extreme variant of RAO the "Eve Theory" or "Complete Replacement".[20] Brauer (1989, 1992, 2001) has named his own model Replacement with Hybridization despite not distinguishing this to the standard RAO model: "Brauer also contends that such hybridization probably did not occur and that if it did its effects were negligible".[21] Aiello (1993) made the mistake of treating RAO and Brauer's model as distinct, writing the latter "allows for a greater extent of hybridization", when contrarily "the fact Brauer views hybridization to be basically insignificant is reflected in his recent dismissal of any evidence of continuity in the early modern Europeans from Mladec"[22] and Brauer (2007) "does not see himself as promoting a model that is different from the African replacement [RAO] model".[23]
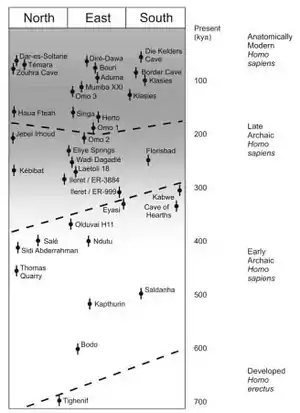
Another misconception is that RAO has to equate African anatomical modernity with a speciation event. Chris Stringer, a leading proponent of RAO notes that "whether we regard the morphological differences between the Neanderthals and modern humans as intraspecific or as specific, we are certainly not dealing with a bio-species concept with absolute fertility boundaries" and that RAO does not "necessarily entail accepting that dispersing modern humans must have represented a new species".[26] This is rather a broader problem in paleontology, since by the nature of fossil evidence, paleontologists are forced to deal with a morpho-species concept rather than a bio-species concept, i.e. paleontologists can never really know whether two similar looking fossils could have produced viable offspring when they were alive.[27]
Criticism of morphological continuity
RAO proponents reject the Multiregional claim there was morphological continuity between archaic humans and early moderns outside of Africa (the period before and after the replacement time). The persistence of Neanderthal skeletal traits in Upper Paleolithic European skulls would demonstrate a significant (non-negligible) degree of archaic genetic admixture (since phenotype is an expression of genes) and falsify the RAO hypothesis, but many studies detect none, e.g. a major study that included informative ancestral non-metrics in the Neanderthal skull found:
“”Of 10 relevant cranial traits examined in that study (Brauer and Broeg, 1998), not a single character indicative of Neanderthal ancestry was found in any of these Upper Paleolithic specimens.[28] |
This denial of morphological continuity does not mean proponents of the Recent African Origin (RAO) hypothesis deny there was some minimal interbreeding between archaic and modern humans, but any genetic admixture is considered too trivial to show up in skeletal/anatomical features.
Morphological traits cited in Multiregionalism and Assimilation model literature as evidence for significant Neanderthal (or other archaic) continuity are instead claimed by RAO proponents to be homoplasies, acquired through convergent evolution: "these features could have continually re-evolved in the same region, producing homoplasy rather than phylogenetic continuity",[29] as cranial traits that are not region-specific/are poorly informative about ancestry (i.e. found at high frequency in multiple regions[30][31]), or as non-homologous features (problematic character states[32]).
“”Opponents of MRE have argued that the traits used to support continuity actually represent shared primitive retentions, characteristics of limited or questionable genetic background or are not unique enough to specific regions to be used as
regional indicators – or some combination of the above (Pearson 2004; Hublin and Bailey 2006; Brauer et al. 2006).[33] |
See also
References
- In the media and among layperson's the RAO hypothesis is more commonly known as the Out of Africa (OOA) theory.
- It is debated where in Africa: "And it is still uncertain whether the pattern of modern human evolution in Africa was essentially continent-wide, or was a more localized phenomenon. African fossils from 100–250,000 years ago show a great deal of anatomical variation, and we are only sampling a tiny part of the whole continent's populations at present (there is no significant record from much of central and western Africa, for example, but we know people were there from lithic evidence). Did the early modern morphology evolve gradually and spread outwards from, say, East Africa, replacing more archaic forms? Or could there have been an African version of multiregionalism, with modern genes, morphology and behaviour coalescing from various populations across the continent?" (Stringer, 2007)
- Prufer et al. 2014.
- Stringer, 2014.
- "The reality is that the actual amount of this contribution to present-day humans is very low and that its phenotypic significance is arguably almost negligible. According to the latest estimates, non-African humans carry around 2% of Neanderthal DNA, with a slightly higher rate in Asia than in Europe." (Hublin, 2014)
- "A common theme among the RAO-supporting commentators (Harvati, Hublin, Stringer) is that the 1%–4% contribution of genes to some modern humans from Neanderthals (and similar genetic contributions from the “Denisovans”) is insignificant, and therefore RAO remains the most viable model of modern human origins." (Holliday et al. 2014)
- Fu et al. 2015.
- "In the past Neandertal genes were more common in Europe than they are today. This is in line with various contentions that many if not most Neolithic populations in Europe came from western Asia. European Neolithic populations were largely a consequence of population replacement and subsequent expansion: some 70 percent of the Neolithic settlements are of Levantine origin (Chihki et al. 2002), Neolithic hunter-gatherers of northern Europe have a genetic profile that is not often found in living populations of the region, continued Neolithic gene flow between farmers from western Asia and local and hunter-gatherer populations created the current pattern of genetic variation (Skoglund et al. 2012)." (Wolpoff & Caspari, 2013)
- Smith & Ahern, 2013.
- "I concur with Holliday et al. (2014) that Assimilation has provided, and continues to provide, a more complete and more operative working model of modern human emergence." (Zilhao, 2014)
- Ahern et al. 2013.
- Rogers & Bohlender, 2015.
- Fu et al. 2014.
- Stringer, 1989.
- Stringer, 1992.
- "We recognize that such hybridization might have occurred, but is of minor or negligible significance for the ancestry of living humans." (Stringer & Brauer, 1994)
- "Stringer (1992) has clarified his stance to allow for some hybridization between anatomically modern populations and local archaic groups, but states that the evidence for this has not been established, and that any such non-conspecific interbreeding would not have led to significant contributions to the modern gene pool by the archaic populations. Both Cann (Cann, Rickards & Lum, this volume; cf. Cann, Stoneking & Wilson, 1987) and Stringer (pers. comm.) now discuss hybridization as a distinct possibility, yet they continue to believe that the contributions of archaic populations were insignificant and unlikely to have been substained for more than a few generations." (Simmons, 1994)
- "Stringer (1992), Cann (1992), and other out-of-Africa proponents (e.g., Bräuer, 1989) have stated that limited gene flow between modern humans and Eurasian archaics may have occurred but that this would not be inconsistent with the out-of-Africa model." (Ahern et al. 2013)
- "While RAO has accepted that some 'hybridization' may have occurred between Neanderthals and early moderns, it is clear that the extent of the Neandertal genetic input is considered essentially insignificant (e.g. Stringer and Brauer, 1994; Brauer and Stringer, 1997; Brauer and Broeg, 1998; Klein, 1999; Brauer, 2001)." (Smith et al. 2005)
- Brauer & Stringer, 1997.
- Cartmill & Smith, 2009: 473.
- Smith, 2002.
- Collard & Dembo, 2013.
- Brauer, 2008.
- Mbua and Brauer (2012) write: "a sequence of three evolutionary grades or morphs within one biological species H. sapiens appears in agreement with the current evidence for Africa". They consider the species Homo sapiens to be 500,000-600,000 years old, and date modern humans (as a grade) 200,000 BP, and that "changes towards modern anatomy are most clearly demonstrable for eastern Africa".
- Brauer & Stringer, 1997.
- Werning, 2013 PLOS Blogs.
- Brauer et al. 2006.
- Stringer & Brauer, 1994.
- "One of us (Stringer, 1992) also checked late Pleistocene north African cranial samples with regard to the occurrence of supposed 'mongoloid' and 'australoid', regional features. It became evident that relevant Chinese features, such as transversely flat faces and shoveled upper incisors, and southeast Asian traits, such as high prognathism, strong supraorbital region, everted malars, and malar tuberosities, all occur with moderate or high frequencies among these North Africans as well." (Brauer & Stringer, 1997)
- "The regional features claimed by the Multiregional Model to characterize the East Asian and Australian evolutionary lines are not exclusive to these regions." (Lahr, 1994)
- An example is the occipital bun found in Neanderthals and early modern Europeans. Yet, the projections of the occipital bone are not homologous (of the same structure, position): "In early modern Europeans, the occipital projections are not only placed 'in the context of a rather different cranial shape' compared to that of Neanderthals (Churchill and Smith. 2000) but they also differ in a number of special details such as the extent of lambdoidal flattening, the angulation between the nuchal and occipital planes, and the height of the vertical face of the posterior occiput (Caspari, 1991). These differences led Smith (1982) to suggest the term 'hemibun' for the occipital projections in Upper Paleolithic specimens." (Brauer, 2007)
- Smith, 2010.
Sources
- Ahern, J. C. M., Janković, I., Voisin, J., Smith, F. H. (2013). "Modern Human Origins in Central Europe". In: Smith, F. H & Ahern, J. C. M. (eds.). The Origins of Modern Humans: Biology Reconsidered. New York: Wiley.
- Aiello, L. C. (1993). "The fossil evidence for modern human origins in Africa: a revised view". American Anthropologist. 95(1): 73–96.
- Brauer, G. (1989). "The evolution of Modern Humans: A comparison of the African and non African evidence". In: Mellars, P. A. & Stringer, C (eds.). The Human Revolution. Edinburgh University Press.
- Brauer, G. (1992). "Africa's place in the evolution of Homo sapiens". In: Brauer G. & Smith, F. H. (eds.). Continuity or Replacement: Controversies in the Evolution of Homo sapiens. Balkema.
- Brauer, G., Stringer, C. (1997). "Models, polarization and perspectives on modern human origins". In: Clark, G. & Willermett, K. (eds.). Conceptual Issues in Modern Human Origins Research. New York: Aldine deGruyter.
- Brauer, G., Broeg, H. (1998). "On the degree of Neandertal-modern continuity in the earliest Upper Paleolithic crania from the Czech Republic: evidence from non-metrical features". In: Omoto, K., Tobias, P. (eds.). The Origins of Past Modern Humans: Toward Reconciliation. Singapore: World Scientific.
- Brauer, G. (2001). "The 'out-of-Africa' model and the question of regional continuity". In: Tobias P.V., Raath M.A., Moggi-Cecchi, J., Doyle G. A. (eds.). Humanity from African naissance to coming millennia. Firenze, Florence & Witwatersrand University Press.
- Brauer, G., Collard, M., Stringer, C. (2004). "On the reliability of recent tests of the Out of Africa hypothesis for modern human origins". Anat. Rec. A. Discov. Mol. Cell. Evol. Biol. 279(2): 701-707.
- Brauer, G., Broeg, H., Stringer, C. (2006). "Earliest Upper Paleolithic crania from Mladec, Czech Republic, and the question of Neanderthal-modern continuity: metrical evidence from the fronto-facial region". In: Havarti, K & Harrison, T. (eds.). Neanderthals Revisited. Dordrecht: Springer.
- Brauer, G. (2007). "Origin of modern humans". In: Henke, W., Tattersall, I. (eds.). Handbook of Paleoanthropology (vol. 3). New York: Springer.
- Brauer, G. (2008). "The origin of modern anatomy: by speciation or intraspecific evolution?". Evolutionary Anthropology. 17(1): 22-37.
- Brauer, G. (2012). "Middle Pleistocene diversity in Africa and the origin of modern humans". In: Hublin, J. J & McPherron, S.P. (eds.). Modern Human Origins: A North African Perspective. Springer.
- Brauer, G., Stringer, C. (1997). "Models, polarization and perspectives on modern human origins". In: Clark, G. & Willermett, K. (eds.). Conceptual Issues in Modern Human Origins Research. New York: Aldine deGruyter.
- Cann, R. L., Stoneking, M., Wilson, A. C. (1987). "Mitochondrial DNA and human evolution". Nature. 325: 31-36.
- Cartmill, M., Smith, F. H. (2009). The Human Lineage. Wiley-Blackwell.
- Collard, M., Dembo, M. (2013). "Modern Human Origins". In: A Companion to Paleoanthropology. Begun, David R. (ed.). Wiley-Blackwell.
- Holliday, T. W., Gautney, J. R., Friedl, L. (2014). "Right for the Wrong Reasons: Reflections on Modern Human Origins in the Post-Neanderthal Genome Era". Current Anthropology. 55(6): 696-724.
- Mbua, E., Brauer, G. (2012). "Patterns of Middle Pleistocene hominin evolution in Africa and the emergence of modern humans". In: Reynolds, S. C & Gallagher, A. (eds.). African Genesis: Perspectives on hominin evolution. Cambridge University Press.
- Prufer, K., Racimo, F., Patterson, N., Jay, F., Sankararaman, S., Sawyer, S., Pääbo, S. (2014). "The complete genome sequence of a Neanderthal from the Altai Mountains". Nature. 505(7481): 43-49.
- Simmons, T. (1994). "Archaic and modern Homo sapiens in the contact zones: evolutionary schematics and model predictions". In: Nitecki, M. H. & Niteck, D. V. (eds.). The Origins of Anatomically Modern Humans. New York: Plenum Press.
- Smith, F. H. (2002). "Migrations, radiations and continuity: Patterns in the evolution of Middle and Late Pleistocene humans". In: Hartwig, W. (ed.). Primate Fossil Record. Cambridge University Press.
- Smith, F. H., Janković, I., Karavanić, I. (2005). "The assimilation model, modern human origins in Europe, and the extinction of Neandertals". Quaternary International. 137(1): 7-19.
- Smith, F. H. (2010). "Species, populations and assimilation in later human evolution". In: A Companion to Biological Anthropology. Larsen, C. (ed.). Wiley-Blackwell.
- Smith, F. H., Ahern, J. C. M. (2013). "Introduction: Thoughts on Modern Human Origins: From 1984 to 2012". In: Smith, F. H & Ahern, J. C. M. (eds.). The Origins of Modern Humans: Biology Reconsidered. New York: Wiley.
- Stringer, C. (1989). "Documenting the origin of modern humans". In: E. Trinkaus (ed.). The Emergence of Modern Humans. Cambridge University Press.
- Stringer, C. (1992)."Replacement, Continuity and the Origin of Homo sapiens". In: Brauer G. & Smith, F. H. (eds.). Continuity or Replacement: Controversies in the Evolution of Homo sapiens. Balkema.
- Stringer, C., Brauer, G. (1994). "Models, misreading and bias". American Anthropologist. 96(2): 416-424.
- Stringer, C. (2014). "Why we are not all multiregionalists now". Trends in Ecology & Evolution. 29(5): 248–251.
- Wolpoff, M., Caspari, R. (2013). "Paleoanthropology and Race". In: A Companion to Paleoanthropology. Begun, David R. (ed.). Wiley-Blackwell.
- Zilhao, J. (2014). "Comment". In: Holliday, T. W., Gautney, J. R., Friedl, L. "Right for the Wrong Reasons: Reflections on Modern Human Origins in the Post-Neanderthal Genome Era". Current Anthropology. 55(6): 715-716.