Common descent
Common descent is the scientific theory that all living organisms on Earth descend from a common ancestor. Although evolution had been suggested as far back as Ancient Greece, the first proponent of common descent appears to have been Erasmus Darwin, the grandfather of Charles Darwin. In his book Zoonomia, published in 1796, he suggested that "all warm-blooded animals have arisen from one living filament". Biologists have evidence that all life developed from a common ancestor which lived just under 4 billion years ago, and virtually all scientists working in the field accept the concept. The structures and functions of all living organisms are encoded in the same basic nucleic molecules, DNA and RNA. Similarities in amino acid sequences between various organisms also suggest common descent, and the fossil record also shows cases in which one plant or animal type evolved into different types over time. The details of this process are shown in clade diagrams.
| We're all Homo here Evolution |
| Relevant Hominids |
| A Gradual Science |
| Plain Monkey Business |
v - t - e |
Creationists reject common descent as it implies an evolutionary model. Young Earth creationists, as Biblical literalists, reject it outright, believing that life was created within a few days less than ten thousand years ago. Intelligent-design proponents may accept some aspects of common descent, as long as they are allowed to claim that a "designer" interfered with the genes and mutations along the way, then deferring what's not explained to "we don't know" what happened.[1][2] Cdesign proponentsists probably believe a muddled overlap of these positions.
Overview of evidence
The most powerful evidence for common descent includes:
- Anatomical homologies — Throughout the domains of life, organisms show a distinct pattern of constraints based on homology in development and construction of the body. For example, tetrapods have five digits because the ancestor of tetrapods had five digits. When a tetrapod does not seem to have five obvious digits, a review of their development shows that they start development with five and that they fuse together later to form fewer numbers.
- DNA and RNA code — Almost all organisms use the same three-letter code for translating RNA into proteins. There are variations, such as the code used by mitochondria and some bacteria and fungi, but the differences are only minor. Regardless of the slight differences, all organisms use the same coding mechanism for translating the code into amino acid sequences.
- Endogenous retroviral insertions — Ancient retroviruses inserted inactivated viral genes into genomes. For a retrovirus to be inherited in all members of a species, a series of highly improbable events must occur. The virus must insert into a gamete cell and it must mutate so it is inactive. That gamete cell must be used to make an embryo that lives to reproduce and whose genome fixates into the population at random location in the genome. This rare event is usually species specific.
- Pseudogenes — Shared errors are a powerful argument for a common source. If two books describe the same concept in similar language, it's possible they just both converged on the same wording. However, if they both share the same grammar or spelling errors it becomes improbable to assume that they did not derive from a common source. There are genes that no longer code for a protein due to a mutation or error. Species often share the same pseudogene with the same inactivating mutation. A famous example of this is the L-gulonolactone oxidase that synthesizes vitamin C. All simians including humans share one pseudogene of inactivated L-gulonolactone oxidase, but the guinea pig has a different pseudogene indicating a different mutation.
- Embryology — The pharyngula stage of embryonic development appears to be highly conserved. At this stage, it is difficult to tell the difference between various vertebrate species. This conserved state screams common ancestry, and the field of evolutionary development has expanded our knowledge of developmental genes and their consequent embryo ontogeny to amazing levels of detail, all thanks to acknowledging common descent.
- Chromosome fusion — Gene fusion or chromosome fusion is when two chromosomes are spliced together. As an example, chimpanzees have one more chromosome than humans do. If the two species share a common ancestor, scientists should be able to figure out what happened to that chromosome. Researchers have found that chromosome 2 in humans is actually the fusion of two separate chimpanzee chromosomes. At the end of each chromosome is a marker called a telomere, which usually appears only on the ends. In human chromosome 2 it also appears in the center, marking where the two ends fused.
- Convergence — The phylogenetic trees constructed using anatomical homology, DNA homology, pseudogenes, endogenous retroviral insertions, and many other methods all converge on a similar looking tree. There are slight differences but the general relationships of the trees are intact. If any of these methods were flawed, they would not converge on the same tree.
- Uniqueness — The complex, predictive patterns of similarities and differences in the world of life have have a unique known explanation — no one has even hypothesized an alternative account for the patterns exhibited — either there is common descent or there is something which is somehow simulating common descent.
Anatomical homology
Anatomical homology refers to the parts of different species that look the same, even when the part performs different functions. These are particularly obvious when the outside is stripped away and the skeleton is examined. For instance, when the skeletal composition of multiple mammals is examined, it is clear that each share many common features. Human arms (ordinarily used for holding, not moving) have similar bone structures to dog forelegs (for walking and running), whale flippers (for swimming) and bat wings (for flying); specifically, two bones that make up the forearm section but a single bone above the elbow. Five fingers are also a common feature across many mammals and related animals.[4] Other examples are abundant as anatomy was paramount in the development of the theory of evolution.
Deep homology
A deep homology is a homology which spans a wide stretch of the tree of life, often not apparent except through study of the proteins or genes involved.[5] One such example is the deep homology between the cluster of genes responsible for the building of blood vessels in vertebrates and in a yeast, for fixing cell walls.[6][7]
Deep homologies are evidence of common ancestry across wide ranges of the tree of life. Furthermore, they are indications of how "irreducibly complex" structures can evolve new functions from precursor structures. And certain cases suggest research programs into problems of medicine (in the vertebrate-yeast case, for the treatment of cancer).
Do fossils validate common descent?
Although fossils are usually not direct evidence in favour of common descent, a purely anatomical resemblance to living species is a form of strong supportive evidence. Some more recent fossils contain traces of tissue or DNA that can be used to determine in what way the fossil is related to living species.[8] As mentioned below, this offers direct evidence for evolution.
Transitional forms
While creationists often claim that no transitional forms have been found, numerous transitional forms have been found over the years.[9][10][11][12]
Of course, one could always move the goalposts and keep asking for more transitional forms between other transitional forms. However, fossilization is the exception to the rule (usually, cadavers decay completely) and seismic, volcanic, tectonic and human activity have destroyed or buried many fossils.
"Transitional forms" is a bit of a misnomer anyway. A case can be made for all forms being transitional or none. All forms alive today seem to be very well adapted to their environment, and none looks obviously "transitional". The same would certainly have been valid in the past; only with the benefit of hindsight can they be labeled "transitional". The only forms that are not obviously "transitional" are those with no living descendants or those that have survived to this day without change. To put it another way, evolution does not occur in between periods in which no changes occur; though it may be slow with occasional bursts in speed, mutation and natural selection are ongoing processes.
RNA / DNA code
All life significantly shares the genetic code based on the molecule DNA and its related molecule RNA. The translation between DNA and RNA codons (groups of three bases) and the corresponding amino acids is almost the same in all known forms of DNA-based life on earth, from humans to bacteria. If the translation between DNA, RNA, and amino acid differs, it allows scientists to probe how creatures have evolved and how different taxonomic groups branched apart.
Endogenous retroviral insertions
What is a retrovirus?
A retrovirus is a virus in the family Retroviridae. Different families of viruses carry their genetic information differently: DNA, double-stranded RNA, and single-stranded RNA are all possible. Retroviruses contain their information in RNA, but unlike other RNA viruses, they use a protein called reverse transcriptase to transcribe their RNA into DNA upon entering the host cell, and then insert the DNA copy into the host genome. Once inserted, viral proteins can be transcribed from the viral DNA, though it can also lie dormant for a period. The retrovirus has significant clinical importance since HIV, which causes AIDS, is a retrovirus.
A retrovirus virion (the physical virus, before entry) consists of a lipid envelope, protein capsid, RNA genome, and the reverse transcriptase. Since RNA is never coded back into DNA in an uninfected cell and there is no endogenous reverse transcriptase in the cytoplasm, the retrovirus must bring it along when leaving the host cell. The discovery of reverse transcriptase in retroviruses has been a major boon for genetic research, allowing for libraries of RNA to be coded into DNA (which is more stable and easier to work with), among other things, as well as in the biomedical industry in which conversion of RNA to DNA is used in many genetic treatments and research.
Endogenous retrovirus
Reverse transcriptase is prone to making errors in transcription. Sometimes these errors will inactivate the retrovirus's genes and the host cell will not produce new viruses. These mutated strands of retrovirus DNA are still integrated in the host genome. In some cases in a multicellular organism, this will be in a germline cell (i.e., egg or sperm). This means that any offspring generated from that germline cell will have the inactivated retrovirus gene in its genome. These are referred to as endogenous retroviral insertions (ERVs), and it has been estimated that up to 10% of the human genome consists of these types of inserts.
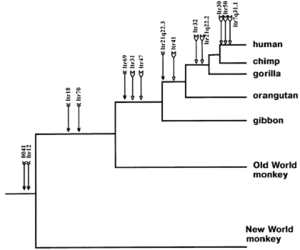
ERVs are usually species-specific, inserted almost randomly in the host genome, and the error or mutation that inactivated the gene is random. If two organisms share the same ERV in the same location with the same inactivation mutations, then they almost certainly share them due to common inheritance and not two separate infections. Researchers analyze shared ERV insertions across species to construct phylogenetic trees.[13] For example, the common ERVs in simians indicates they share a common genome. When phylogenetic trees are constructed based on the pattern of ERVs, they indicate humans share more ERVs with chimps than either share with gorillas. Other examples are known.[14][15] This is strong evidence for common descent.
Pseudogenes and shared errors
What is a pseudogene?
Pseudogenes are genes present in an organism's genome that have lost the ability to code for proteins due to mutation.[16] They were first identified and dubbed in the late 1970s when researchers began finding non-coding regions in some organisms that were similar to actual coding genes in other organisms.[17] So far an estimated 19,000 pseudogenes have been identified in the human genome, almost equal to the total number of coding genes (21,000).[17] Humans have many pseudogenes including L-gulonolactone oxidase, which is used to synthesize vitamin C. Research reports that this gene was inactivated in the common ancestor of all simians.[18]
Pseudogenes have been identified in a wide range of organisms from bacteria to mice to humans. The total number of pseudogenes in a given genome is not predictable, but specific pseudogenes are often compared across species to elucidate complex evolutionary relationships.[17]
Pseudogenes are often difficult to parse from the large number of non-coding base pairs in the genome. Convention requires two elements to be present to label a sequence a pseudogene. The first is homology, which is the requirement that a sequence be demonstrated to descend from a functional copy of the gene. The second is non-functionality, which is the requirement that the gene not code for a protein in the organism in question.[17]
Since all pseudogenes are hypothesized to be descended from a parent functioning gene, the first step is to identify the parent gene. Computer programs to compare sequences of DNA across species are used to accomplish this.[17] Using the phylogenetic relationships between species, one can decrease the search time by looking at species that share a more recent common ancestor.[19] Once a functioning copy of a gene is detected, its sequence is compared to the pseudogene. A high correlation in base pairs is used to assign homology. Non-functionality can be demonstrated by attempting to transcribe the sequence in vitro.[17]
Pseudogenes as shared errors
In copyright law there is a problem when determining if one source has copied another source because it is possible, particularly with topics in narrow subfields, that two authors could converge on a similar sounding passage to describe the same concept. However, errors in the passages are independent of the subject of the text and of each other. Multiple shared errors, particularly in grammar or spelling, become increasingly improbable for two independent writings. If there are several shared errors between two passages the only reasonable explanation is that one is a copy of the other or that both were copied from a common source.
This same concept applies to pseudogenes. While it is possible that two independently-evolved genes will look the same because they both do the same thing, after an error invalidates a gene, the sharing of this same error between two species is extremely strong evidence showing that both species derived from the same source in which the mutation first appeared. By analyzing shared inactivation mutations in non-functional pseudogenes, scientists can construct phylogenetic trees and prove common descent. One such example is hemoglobin; hemoglobin is the protein in red blood cells that transports oxygen to the blood stream. Hemoglobin is made up of four parts called polypeptides, it has two copies of a part of the protein called alpha-globin and the same for beta-globin. The genes for alpha-globin can be found on chromosome #16, and the genes for beta-globin on chromosome #11. There are six sequences for the building of the polypeptides, five of them work, one doesn't. It's what's called a "pseudogene", so it doesn't work, but it's still recognized as a gene because of its similarity with real genes. So the errors in the gene renders the gene defunct, non-functional. What's important about this is because it can be found in three organisms, the gorilla
Vitamin C and our simian common ancestor
One famous shared error that provides strong evidence for a shared ancestry between humans and other simians is gulonolactone oxidase. This enzyme catalyzes the reactions needed to produce ascorbic acid (vitamin C).[21] While the gene used to produce this enzyme is present in most animals, it has been inactivated in others due to mutation. Animals that can no longer synthesize vitamin C include simians, guinea pigs, and several species of fruit bats. Since vitamin C is readily available through the consumption of fruit, the inactivation of this gene was not a significant detriment despite the potential for scurvy. Hence, natural selection did not eliminate the mutated gene.
The mutation that causes inactivation of the L-gulonolactone oxidase gene is different depending on the group the organism is in. All extant guinea pigs share the same inactivation mutation, while all extant simians share a different mutation. The likelihood of two different species sharing the same inactivation mutation is statistically improbable; a better explanation is that the sharing is actually due to common inheritance. The most recent common ancestor of all extant guinea pigs developed one mutation in the gene, while a different mutation developed in the most recent common ancestor in simians.
Biologists have discovered other examples of pseudogenes and have related them to phylogeny.[22][23]
Embryology
Conserved development and the pharyngula
Pharyngula is the term used to describe the phylotypic stage of development in embryology.[24] Taxonomically diverse vertebrate embryos all seem to converge to a very similar morphology.[25] The term was coined in 1981 and stems from the characteristic pharyngeal arches that appear.[25] It is defined as "The point during development when the basic body plan for a particular higher-level taxa is visible and when all the members of this taxon look most similar."[26]
The evidence suggests that this stage represents the basic vertebrate body plan in the common ancestor of all vertebrates,[25] although there is some dispute as to how similar embryos are and to the reality of this stage.[27]
There are six stages to embryonic development, and the pharyngula stage is towards the middle. In the early stages of development there is significant diversity in the morphology of embryos. This diversity decreases over time until the pharyngula stage during which they are most similar (often difficult for anyone but a trained embryologist to differentiate), and finally in the last stages of development morphology diversifies again.[28] It is hypothesized that the reason the pharyngula stage is so morphologically constrained is that this is the point when sequential activation of Hox genes is initiated so any strong deviations from the developmental plan would lead to drastic changes in the final phenotype of the organism.[27]
Hox genes and ontogeny
The Hox genes are a set of regulatory genes common in all life forms that seem to be largely responsible for orchestrating the ontological development of an organism's body plan. Hox genes are not expressed as proteins but rather act to turn on and off expressed genes in the genome to develop specific body parts such as eyestalks or arms. Early in the development of the embryo, patterns of Hox genes will be turned on and off based on the cell's position relative to other cells and the perceived orientation of the embryo (up/down, left/right). This pattern of activated Hox genes controls the development of what anatomical features emerge. By activating Hox genes in the lab or moving cells around once the Hox genes are activated significant changes occur in morphology (such as eyestalks instead of legs).[29]
Hox genes are fascinating to study in all aspects of biology, but they also offer several strong lines of evidence for common descent. A striking element of Hox genes is that they are highly conserved. An embryologist can knock out a Hox gene in a fly, replace it with a similar one from an earthworm, and still wind up with a normally developed fly. Because the Hox genes are so important to the development of an organism, there is little room for beneficial mutation and natural selection acts to conserve the basic structure. (It could be argued this is a product of a "common designer", although there is no parallel in the design of synthetic objects). What really makes this powerful evidence for common descent is the pattern of the development of new Hox genes.
New Hox genes arise from duplication of old Hox genes. One of the Hox genes is redundant and hence susceptible to evolutionary changes. Researchers are able to take Hox genes that are found in more derived organisms that are not found in more ancestral organisms and show how the new Hox genes are slightly modified Hox genes from evolutionary older organisms. It is then possible to construct phylogenetic trees for the actual Hox genes. The amazing part is that this phylogenetic tree for the Hox genes matches phylogenetic trees for the organisms themselves. This only makes sense from the viewpoint of common descent.

Chromosome fusion
Another piece of evidence that makes clear the factual reality of common descent is chromosome fusion that occurs across a wide variety of species in different ways.[30] Fusion of chromosomes decreases the chromosome numbers in a descendant species. (Alternatively, a split in a chromosome increases the chromosome number.) The pattern of these fusion events generates characteristic phylogenetic trees offering proof of common descent.
One famous example is a fusion event that indicates the evolution of Homo sapiens from a common ancestor of humans and chimpanzees.[31] While all other great apes have 24 pairs of chromosomes, humans have 23 pairs of chromosomes. Human chromosome 2 looks almost identical to two of the chimpanzee's chromosomes stacked on one on top of the other, indicating chromosome fusion.[32]
Chromosomes form light and dark bands on a karyotype that can be compared to see how similar they are. The light and dark banding patterns of the two chimp chromosomes match that of the single human one. Also, on the end of each chromosome are a series of repetitive DNA sequences called telomeres. These sequences are found typically only on the tips of the chromosome. For chromosome 2, scientists find them in the middle, as if the tips of two chromosomes merged together.
In addition, chromosomes have a centromere which link the two sister chromatids together. While there are usually only one of these distinct regions on a chromosome, there are two centromeres on chromosome 2. One is "non-functional", but its genetic code matches the other centromere from the second chimpanzee chromosome.
Clearly from this evidence human chromosome 2 is a fusion event between two chromosomes found in chimpanzees in distinct form. This means that humans inherited these chromosomes from a common ancestor with chimpanzees.
Biologists have discovered other gene fusions that are consistent with common descent.[33][34]
Convergence
Convergence is the strongest piece of evidence of all. The most iconic symbol of common descent is the tree of life, a branching structure showing the theorized relationships between all of life, tracing back to the last universal common ancestor. The relationships of various extant and extinct species can be constructed using any evidence discussed above. Before the modern genetic tools, anatomical homologies were used to construct the relationships. After scientists employed genetics in tree construction, several long debates about relationships were settled, and a few surprises and changes in the tree emerged. However, the tree constructed from genetic information was astoundingly similar to that constructed from anatomical homologies. Different genetic tools such as DNA structure, chromosome structure, and endogenous retrovirus insertions can all be used independently to construct individual trees. While there may be slight changes or deviations between trees, all methods converge on similar relationships. This convergence is powerful evidence for the validity of common descent. Each method uses independent observations to produce results that only common descent predicts.
Creationists like to make comparisons to the style changes of synthetic objects as examples of how trees between designed objects could look the same as the tree of life. These examples rely almost exclusively on what would be "anatomical homologies." A Toyota truck and a Ford truck would be placed close on such a tree. However, when you look down to the component level, a Toyota sedan made in the same factory as their trucks would be closer to them than a truck manufactured in Detroit. One would expect this lack of convergence among multiple independent tree constructions if there is no true common descent. However, in life, this convergence between models is exactly what scientists find.
Creationist hubris
Most creationists (and, indeed, most people) would gladly admit they do not understand quantum mechanics and are unqualified to make a judgment about its validity. In contrast, they have no such qualms about evolution and common descent.[35]
While simple on the surface (as taught in the average high school), the theory of evolution in the form of the modern synthesis is as complex as quantum mechanics with a complex interplay of disparate evidence. The interaction between genetic mutations, organism development, and events in the life of an organism is incredibly complex. Since evolution occurs at a population level, the interaction within the population and its interaction with the environment adds another layer of complexity. This contrasts with the gut-level or biblical rejection of the theory. For example, one evolutionary law is Dollo's law of irreversibility, which states that an organism can't "devolve" back into phenotype of the ancestral organism, there are exceptions to this law, so this is a generalization. However, according to creationism, this law shouldn't exist, yet it does. There is a proposed mechanism to this law according to a 2009 study. In the study, they changed a protein structure by way of amino acid substitutions. Said protein was a hormone receptor that was able to bind two proteins together to form a new hormone; when they tried reversing it they found that several mutations would have to occur at once, which is extremely unlikely, thus forming both experimental evidence of Dollo's law and potentially explaining why evolution tends to go in one direction.[36]
See also
- Cladistics
- Ring species
- Baraminology
- Branches of science you have to ignore to believe in young Earth creationism
External links
- See the Wikipedia article on Common descent.
- See the Wikipedia article on Evidence of common descent.
- 29+ Evidences of Common Descent on talk.origins
- Chromosome 2 Fusion and Bayes Theorem: Support for Common Ancestry After All
- Beyond Reasonable Doubt: Evolution from DNA Sequences
- Testing Common Ancestry: It’s All About the Mutations
References
- Jack Krebs Asking About Common Descent posted by Dave S. (February 11, 2006) Uncommon Descent.
- PZ Myers gives an account of different IDers agreeing, denying or not taking a stance on common descent.
- Gegenbaur, Carl Grundzüge der vergleichenden Anatomie. 2. umgearb. Auflage. Mit 319 Holzschnitten. Leipzig, Verl. von Wilhelm Engelmann, 1870. 892 pp. fig. 223. p. 692.
- Taxonomy: Classifying Life by John W. Kimball (16 December 2013) Kimball's Biology Pages
- Neil Shubin. C. Tabin, Sean B. Carroll, "Fossils, genes and the evolution of animal limbs", Nature, volume 388, issue 6643 (August 1997), pages 639–648, pmid 9262397, doi:10.1038/41710
- Zimmer, Carl. "The Search for Genes Leads to Unexpected Places", The New York Times, New York, April 26, 2010.
- Kriston L. McGary, Tae Joo Park, John O. Woods, Hye Ji Cha, John B. Wallingford, and Edward M. Marcotte, "Systematic discovery of nonobvious human disease models through orthologous phenotypes", Proceedings of the National Academy of Sciences, volume 107 number 14 (2010), pages 6544-6549, doi:10.1073/pnas.0910200107,
- Neandertal DNA Mark Rose, Archaeological Institute of America
- Fins to Limbs: New Fossil Gives Evolution Insight National Geographic, April 1, 2004
- Transitional forms Evolution Library, University Of Berkeley
- Human Ancestry: Species ArchaeologyInfo.com
- Transitions: The Evolution of Life
- 29+ Evidences for Macroevolution. Part 4: The Molecular Sequence Evidence by Douglas Theobald (2004) Talk Origins.
- An example in insects is found here.
- An example in birds is found here.
- Petrov, D.A, Hartl, D.L. (2000). Pseudogene evolution and natural selection for a compact genome. The American Genetic Association 91:221-227.
- Gerstein, M, Zheng, D. (2006). The real life of pseudogenes. Scientific American 95:48-55.
- Eating What Comes Naturally: An Examination of Some Differences Between the Dietary Components of Humans and Wild Primates by Katharine Milton. Talk prepared for: The Origins and Evolution of Human Diet, 14th International Congress of Anthropological and Ethnological Sciences, July 26-August 1, 1998, Williamburg, Virginia, USA. (archived from December 6, 2006).
- Bensasson, D., Zhang, D., Hartl, D., Hewitt, G. (2001). Mitochondrial pseudogense: evolution's misplaced witness. Trends in Ecology and Evolution 16: 314-321.
- Kitzmiller v. Dover: Kenneth Miller Direct Examination Day One, AM
- "Molecular basis for the deficiency in humans of gulonolactone oxidase, a key enzyme for ascorbic acid biosynthesis", American Journal of Clinical Nutrition, Vol 54, 1203S-1208S
- Phylogenetic Analysis and Identification of Pseudogenes Reveal a Progressive Loss of Zona Pellucida Genes During Evolution of Vertebrates by Ghylène Goudet et al. (2008) Biology of Reproduction 78(5):796-806. DOI:10.1095/biolreprod.107.064568
- Phylogeny and Classification of Birds: A Study in Molecular Evolution
- Pharyngual Period
- Irie, N., Sehara-Fujisawa, S. (2007). The vertebrate phylotypic stage and an early bilaterian-related stage in mouse embryogenesis defined by genomic information. BMC Biology 5:1.
- Slack, J. M. W., P. W. H. Holland, and C. F. Graham. 1993. The zootype and the phylotypic stage. Nature 361:490–492.
- Callazo, A. (2000). Developmental variation, homology and the pharyngula stage. Systematic Biology 49:1, 3-18.
- Embryo Stages
- Pharyngula — The Hox Code
- Changes in chromosome number during evolution Talk Origins archive.
- MacAndrew, Alec Human Chromosome 2 is a fusion of two ancestral chromosomes. From evolution pages.
- IJdo JW, Baldini A, Ward DC, Reeders ST, Wells RA, Origin of human chromosome 2: an ancestral telomere-telomere fusion. Proc Natl Acad Sci U S A 1991 Oct 15;88(20):9051-5. Available here
- Gene fusion is a possible mechanism underlying the evolution of STA1 by I. Yamashita et al. (1987) J. Bacteriol. vol. 169 no. 5 2142-2149. doi:10.1128/jb.169.5.2142-2149.1987
- Evolution of Multi-domain Proteins by Gene Fusion and Fission by Sarah K. Kummerfeld et al. (2004) ISMB.
- Cases of opponents totally missunderstanding evolution are common, just one example is Bob Enyart of "Colorado Right to Life" who imagines evolutionary theory involves humans evolving from pigs and dogs as well as apes. See Colorado birth control scheme causes drop in teen pregnancy
- Bridgham, Jamie T.; Eric A. Ortlund; Joseph W. Thornton (2009). "An epistatic ratchet constrains the direction of glucocorticoid receptor evolution". Nature. 461 (7263): 515–519. ISSN 0028-0836. PMID 19779450. doi:10.1038/nature08249.